UNIVERSE
made of an infinite amount of space,
matter and time.
the start of the universe, the universe
has no beginning and no end; perhaps
the same photons that have always been
in the universe continue to move in the
space that has always been.
particles of light humans have named
"photons". Photons are the base unit
of all matter from the tiniest
particles to the largest galaxies.
meters every second in a line, but as
pieces of matter, can be slightly
slowed from the force of gravity, and
stop for an instant when they collide.
and so photons form structures such as
protons, atoms, molecules, molecule
groups (like all of life of earth),
planets, stars, galaxies, and clusters
of galaxies.
galaxies we can see are only a tiny
part of the universe. Most of the
galaxies in the universe we will never
see because they are too far away for
even 1 particle of light from them to
be going in the exact direction of our
tiny location, or are captured by atoms
between here and there.
clear. Photons form gas clouds of
Hydrogen and Helium, these gas clouds,
called nebuli condense to form galaxies
of stars. The stars emit photons back
out into the rest of the universe,
where they collect and form clouds
again. Around each star are many
planets and pieces of matter. On many
of those planets intelligent life
evolves. This life moves their stars
out of spiral galaxies to form globular
clusters, and ultimately to transform
spiral galaxies into elliptical
galaxies that travel the universe
looking for more matter to fuel their
movement.
It may very well be that stars at this
scale are photons, spiral galaxies
charged particles, globular galaxies
neutral particles, and galactic
clusters atoms at a much larger scale
in an infinite macro and micro scale.
from a gas cloud that formed by
capturing matter in the form of light
from other stars, from the remains of a
previously destroyed galaxy, or some
combination of the two.
eventually orbit forms, perhaps in a
nebula, when matter in the nebula
starts accumulating and rotating as a
result of gravity, or from the remains
of an exploded star that condensed
again under the influence of gravity.
move closer to the center and lighter
atoms are sent farther out.
Terrestrial planets are red hot, have
surface of melted rock, all lighter
atoms float to the surface of the
molten planets. All the H2O from the
first earth oceans and lakes is in the
atmosphere in gas form.
ways:
1) spherical planet collides with
earth, moon forms from remaining matter
in ring around earth.
2) spherical planet is
caught in earth orbit
3) moon of earth forms
naturally from original matter of star
system in orbit around earth.
4,571 million years old.

[1] The ''Zag'' meteorite fell to Earth
in 1988 COPYRIGHTED
source: http://news.bbc.co.uk/1/hi/sci/t
ech/783048.stm
Apollo missions (4.53 billions old).

[1]
http://www.nasm.si.edu/exhibitions/attm/
atmimages/S73-15446.f.jpg
http://www.nasm.si.edu/exhibitions/attm/
nojs/wl.br.1.html
source:
LIFE
cools into thin crust, H2O condenses
from the atmosphere by raining, filling
the lowest parts of land to make the
first earth oceans, lakes, and rivers.
meteorite) zircon yet found on earth,
4.404 billion years old, from Gneiss in
West Australia, is evidence that the
crust and liquid water were on the
surface of earth 4.4 billion years
before now.

[1]
http://www.geology.wisc.edu/zircon/Earli
est%20Piece/Images/8.jpg
source:
sugars, the components of living
objects are created on earth. These
molecules are made in the oceans, fresh
water, and or atmosphere of earth (or
other planets) by lightning, photons
with ultraviolet frequency from the
star, or ocean floor volcanos.
nucleotides), proteins (polymers made
of amino acids), carbohydrates
(polymers made of sugars) and lipids
(glycerol attached to fatty acids)
evolved is not clearly known.
Some proteins and nucleic acids have
been formed in labs by using clay which
can dehydrate and which provides long
linear crystal structures to build
proteins and nucleic acids on. Amino
acids join together to form
polypeptides when an H2O molecule is
formed from a Hydrogen (H) on 1 amino
acid and a hydroxyl (OH) on the second.
Are all proteins, carbohydrates, lipids
and DNA the products of living objects?
Is RNA the only molecule of these that
was made without the help of living
objects?
The most popular theory now has RNA
(and potentially lipids) evolving first
before any living objects.
There is still a large amount of
experiment, exploration and education
that needs to be done to understand the
origins of living objects on planet
earth. My opinion is that as soon as
there was liquid water on the earth,
4.4 billion years before now, as zircon
crystals show, the construction of
living objects started on earth.
Perhaps RNA molecules, called
"ribozymes" evolved which can make
copies of RNA, by connecting free
floating nucleotides that match a
nucleotide on the same or a different
RNA, without any proteins. But until
such ribozyme RNA molecules are found,
the only molecule known to copy nucleic
acids are proteins called polymerases.
If such ribozymes exist, then one of
the first coded instructions on the RNA
molecule that was the ancestor of every
living species, must have been the code
to make this ribozyme.
creation of various Transfer RNA (tRNA)
molecules.
Random mutations in the copying (and
perhaps even in the natural formation)
of RNA molecules probably created a
number of the necessary tRNAs (transfer
RNA, an RNA molecule responsible for
matching free floating amino acid
molecules to 3 nucleotide sequences on
other RNA molecules).
This would be a precellular protein
assembly system, where tRNA (transfer
RNA) molecules can build polypeptide
chains of amino acids by linking
directly to other RNA strands.
Part of each tRNA molecule bonds with a
specific amino acid, and a 3 nucleotide
sequence from a different part of the
tRNA molecule bonds with the opposite
matching 3 nucleotide sequence on an
(m)RNA molecule.
Since there are tRNA molecules for each
amino acid (although some tRNAs can
attach to more than one amino acid?),
there must have been a slow
accumulation of various tRNA molecules
for each of the 20 amino acids used in
constructing polypeptides in cells
living now. Perhaps after the
evolution of the first tRNA, the first
polypeptides were chains of all the
same one amino acid. With the
evolution of a second tRNA polypeptides
would have more variety because now two
amino acids would be available to build
polypeptides.
This polypeptide assembly system may
exist freely in water, or within a
liposome. This sytem builds many more
proteins than would be built without
such a system. The mRNA with the code
to make copier RNA, now also contains
the code to produce various tRNA
molecules. These molecules function as
a unit, and proto-cell, with the rest
of the mRNA initially containing random
codes for random proteins.
For the first time, RNA code represents
a template for other RNA molecules, but
also a template for building proteins
with the help of tRNA molecules.
There is some question of where the
origin of the first cell took place,
near volcanos on the ocean floor, or in
fresh water lakes and tidal pools near
volcanos on land, because unprotected
nucleic acids cannot exist for much
time in the ocean because of Sodium and
Chlorine.
Ribosomal RNA moves down mRNA molecules
functioning as a platform for bringing
the mRNA and tRNA molecules together to
assemble polypeptides (proteins).
This rRNA serves as an early ribosome;
objects that serve as sites for
building polypeptides and are found in
every cell. As time continues the
ribosome will grow to include two more
RNA molecules, some protein molecules,
and a second half that will make
polypeptide construction more
efficient.
The rRNA serves the purpose of bringing
amino acids close enough to bond with
each other to form polypeptides.
As an rRNA moves down an mRNA, tRNA
molecules bond with the mRNA and on the
opposite side of the tRNA, a matching
amino acid (separates? from the tRNA
and) attaches to a growing polypeptide
chain.
Now the mRNA that is the
ancestral/progenitor of all of life,
contains the code for the copier RNA,
tRNAs, and the rRNA molecule. These
nucleic acids function as a unit, and
proto-cell.
importance is built, an RNA polymerase.
A molecule that can more efficiently
copy RNA.
is built by the early ribosome protein
making protocell. This protein changes
ribonucleotides into
deoxyribonucleotides. This allows the
first DNA molecule on earth to be
assembled.
Ribonucleotide reductase may be the
molecule that allowed DNA to be the
template for the line of cells that
survived to now.
to copy DNA by assembling DNA
nucleotides from other DNA molecules.
causes the early ribosome to create
reverse transcriptase, a protein that
can assemble DNA molecules from an RNA
molecule template.
With this advance, a DNA molecule can
be constructed that has all of the code
that was stored on the long evolved RNA
molecule. DNA now serves as a more
stable template for making mRNA, each
tRNA, rRNA, and the RNA and DNA
polymerases.
RNA polymerase proteins build RNA
molecules using the new DNA template,
that still perform their original
polypeptide building function together
with the tRNA and rRNA molecules, but
are labeled "mRNA" (Messenger RNA)
because they move from DNA to ribosome.
around DNA, made of proteins. This
membrane holds water inside a cell.
This is the first cell. rRNA
comparison shows that this is most
likely a eubacterium.
DNA produces instructions for
cytoplasm, the cytoplasm is assembled
from proteins made by the ribosome.
For the first time, DNA and ribosomes
are building cell structure. The
templates for each tRNA, rRNA, mRNA and
DNA polymerase proteins are already
coded in a central strand of DNA. DNA
protected by cytoplasm is more likely
to survive and copy. This cell is
heterotrophic and has no metabolism to
produce ATP. Amino acids, nucleotides,
H2O, and other molecules enter and exit
the cytoplasm only because of a
difference in concentration from inside
and outside the cell (passive
transport) and represent the beginnings
of the first digestive system. This
either happens in fresh water lakes or
in salty oceans, perhaps near lava
vents on or under the ocean floor. As
this line of DNA continues to make
copies of itself, all copies now have
cytoplasm. The DNA is composed mainly
of instructions to assemble the nucleic
acids and proteins needed to build
ribosomes, polymerases and cytoplasm.
This cell structure forms the basis of
all future cells of every living object
on earth. These first cells are
anaerobic (do not require free oxygen)
and heterotrophic, meaning that they do
not make their own food: amino acids,
nucleotides, phosphates, and sugars.
These bacteria depend on these
molecules and photons in the form of
heat to reproduce and grow.
A system of division must evolve which
attaches the original and newly
synthesized copy of DNA to the
cytoplasm, so that as the cell grows,
the two copies of DNA can be separated
and the first membraned cells can
divide into two cells. This is the
beginning of the "binary fission"
method of cell division. Division of
the cell begins with the division of
the DNA membrane-attachment site and
separates by the growth of new
cytoplasm.
circle allows the DNA polymerase to
make continuous copies of the cell.
molecules into and out of the cytoplasm
(facilitative diffusion) evolve.
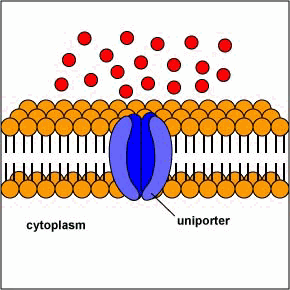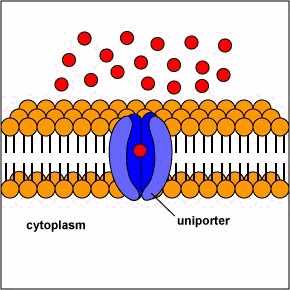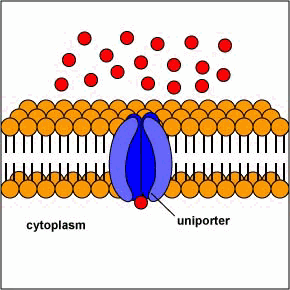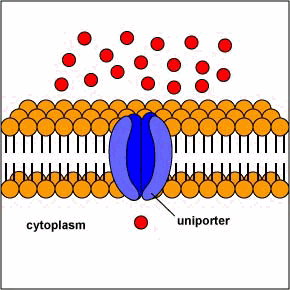
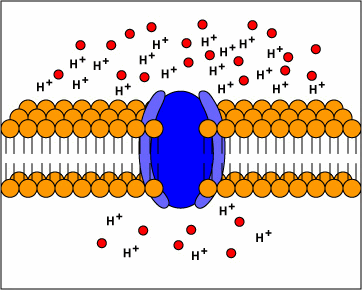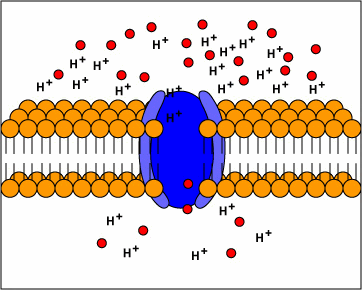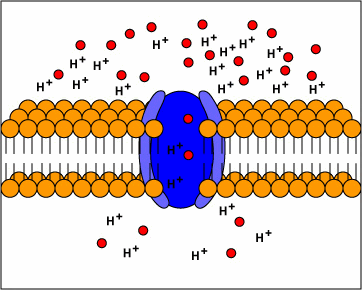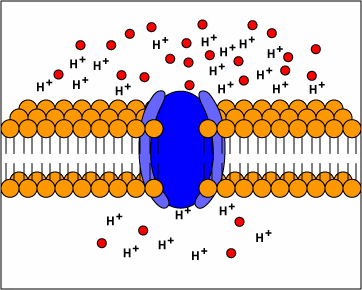
[1] Uniporters are transport proteins
that transport a substance across a
membrane down a concentration gradient
from an area of greater concentration
to lesser concentration. The transport
is powered by the potential energy of a
concentration gradient and does not
require metabolic energy.
source: http://www.cat.cc.md.us/~gkaiser
/biotutorials/eustruct/cmeu.html
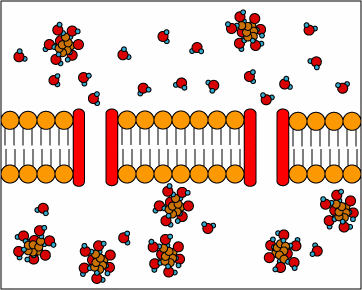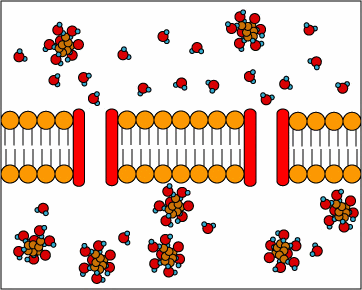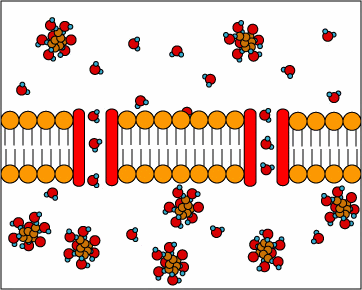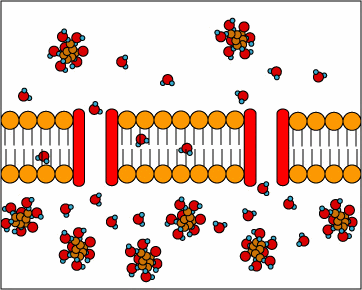
[2] Channel proteins transport water
or certain ions down a concentration
gradient from an area of higher
concentration to an area of lower
concentration. In the case of water,
the channel proteins are called
aquaporins. Water molecules are small
enough that they can also pass between
the phospholipids in the cytoplasmic
membrane by passive diffusion.
source:
from bacteria, or are initially
bacteria. These cells depend on the
DNA duplicating and protein producing
systems of other cells to reproduce
themselves. Over time, more effective,
and efficient virus designs will
survive.
cytoplasm. Cells can now make ATP from
glucose and eventually other
monosaccharides, the end product is
pyruvate.
The glycolysis equation is:
C6H12O6
(glucose) + 2 NAD+ + 2 ADP + 2 P
-----> 2 pyruvic acid, (CH3(C=O)COOH +
2 ATP + 2 NADH + 2 H+
cytoplasm. Cells (all anaerobic) can
now make more ATP and convert pyruvate
(the final product of glycolysis) to
lactate (an ionized form of lactic
acid).
evolves in the cytoplasm. Cells (all
anaerobic) can now convert pyruvate
(the final product of glycolysis) to
ethanol.
that can assemble lipids.
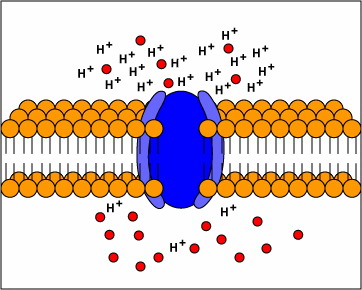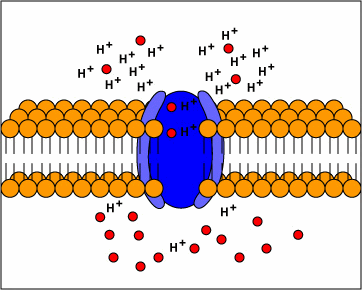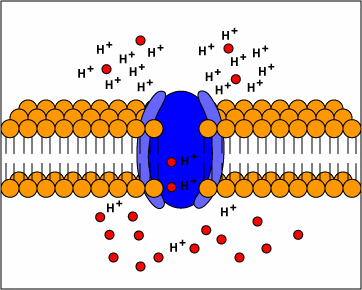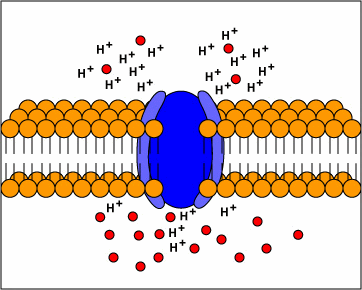
metabolism (ATP) to transport molecules
into and out of the cytoplasm (active
transport) evolve.
[1] TP: not clear what the red circles
are, some kind of molecule I
guess. Antiporters are transport
proteins that simultaneously transport
two substances across the membrane in
opposite directions; one against the
concentration gradient and one with the
concentration gradient. Antiporters
typically use proton motive force to
transport a substrate across the
membrane. The movement of protons
across the membrane (proton motive
force) provides the energy for
transporting the substrate across the
membrane against its concentration
gradient..
source: http://www.cat.cc.md.us/~gkaiser
/biotutorials/eustruct/cmeu.html
[2] Symporters are transport proteins
that simultaneously transport two
substances across the membrane in the
same direction; one against the
concentration gradient and one with the
concentration gradient. Symporters
often use proton motive force to
transport a substrate across the
membrane. The movement of protons
across the membrane (proton motive
force) provides the energy for
transporting the substrate.
source:
evolves in prokaryotes. Now some
prokaryotes can exchange circular
pieces of DNA (plasmids), through tubes
(pili). Conjugation may be the process
that led to sex (cellular fusion) and
also the transition from a circle of
DNA to chromosomes in eukaryotes, since
some protists (cilliates and some
algae) reproduce sexually by
conjugation.

[1] the fertility factor or F factor is
a very large (94,500 bp) circular dsDNA
plasmid; it is generally independent of
the host chromosome. COPYRIGHTED
source: http://www.mun.ca/biochem/course
{kind=link}
s/3107/images/Fplasmidmap.gif

[2] conjugation (via pilus)
COPYRIGHTED EDU
source: http://www.bio.miami.edu/dana/16
{kind=link}
0/conjugation.jpg
allow certain proteins coded by DNA to
not be built, evolve. Proteins bind
with these DNA sequences to stop RNA
polymerase from building mRNA molecules
which would be translated into
proteins. Operons allow a bacterium to
produce certain proteins only when
necessary. Bacteria before now can
only build a constant stream of all
proteins encoded in their DNA.
eubacteria.

[1] This is an image of nitrogen cycle
taken from this [1] EPA website. PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Nitrogen_Cycle.jpg
filment growth evolves in prokaryotes.
prokaryotes evolve. Heterocysts evolve
in cyanobacteria.
Heterocysts are specialized
nitrogen-fixing cells formed by some
filamentous cyanobacteria during
nitrogen starvation.
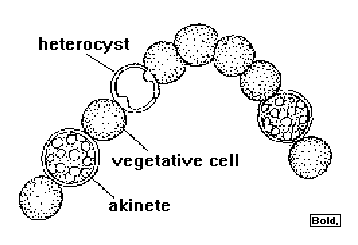
[1] Anabaena COPYRIGHTED EDU
source: http://home.manhattan.edu/~franc
{kind=link}
es.cardillo/plants/monera/anabaena.gif
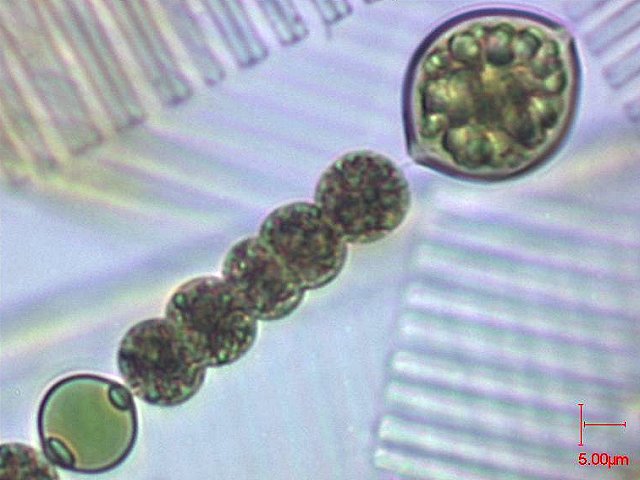
[2] Anabaena smitthi COPYRIGHTED
FRANCE
source: http://www.ac-rennes.fr/pedagogi
{kind=link}
e/svt/photo/microalg/anabaena.jpg
can produce some if not all of their
own food (amino acids, nucleotides,
sugars, phophates, lipids, and
carbohydrates), but require phosphorus,
nitrogen, CO2, water and light in the
form of heat.
There are only 2 kinds of autotrophy:
Lithotrophy and Photosynthesis. These
are lithotrophic cells that change
inorganic (abiotic) molecules into
organic molecules. These cells are
archaebacteria, called methanogens that
perform the reaction: 4H2 + CO2 -> CH4
+ 2H2O. They convert CO2 into Methane.
Methane is better than CO2 for
trapping heat, and could have
contributed to heating the earth.
cells only have Photosystem I.
Photosynthesis Photosystem I evolves in
early anaerobic prokaryote cells. One
of two photosythesis systems,
photosystem I uses a pigment
chlorophyll A, absorbs photons in 700
nm wave lengths best, breaking the bond
betwenn H2 and S. They are anaerobic
and perform the reaction: H2S
(Hydrogen Sulfide) + CO2 + light ->
CH2O (Formaldehyde) + 2S.
evolves in early prokaryote cells.
Photosystem 2 absorbs photons best at
680nm wavelengths, a higher frequency
of light than Photosystem I. These
cells can break the strong Hydrogen
bonds between Hydrogen and Oxygen in
water molecules (more abundant than
Sulphur). This system emits free
Oxygen.
The simple equation of photosynthesis
is: 6 H2O + 6 CO2 + photons = C6H12O6
(glucose) + 6O2. The detailed steps of
photosynthesis are called the "Calvin
Cycle". Prokaryote cells can now
produce their own glucose to store and
be converted to ATP by glycolysis and
fermentation later.
This sytem is the main system
responsible for producing the Oxygen
now in the air of earth.
the "Citric Acid Cycle", and the "Krebs
Cycle") evolves, probably in
cyanobacteria, as a substitute for
fermentaton, by using oxygen to break
down the products of glycolysis,
pyruvic acid, to CO2 and H2O, producing
18 more ATP molecules.
This is the
first aerobic cell, a cell that has an
oxygen based metabolism. This cell
uses oxygen to convert glucose (and
eventually other sugars and fats) into
CO2, H2O and ATP. For example, cells
that oxidize glucose perform the
reaction:
C6H12O6 + 6 O2 + 38 ADP + 38 phosphate
-> 6 CO2 + 6 H2O + 38 ATP
This reaction
(with glycolysis) can produce up to 36
ATP molecules. Cellular respiration is
the opposite (although the specific
reactions differ) of photosynthesis
which starts with H2O and CO2 and
produces glucose.
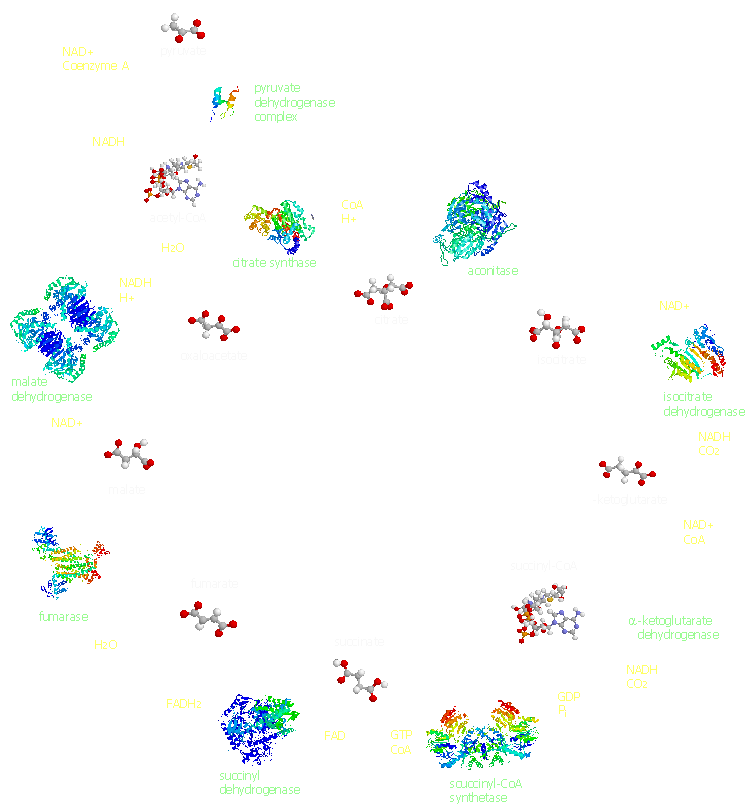
[1] kreb cycle from
http://people.unt.edu/~hds0006/tca/
source:
estimates of when the first Eubacteria
and Archaea evolved. Eubacteria and
Archaea (also called Archaebacteria)
are the two major lines of Prokaryotes.
Prokaryotes are the most primitive
living objects ever found. In contrast
to the later evolved Eukaryotes,
Prokaryotes have a circle of DNA
located in their cytoplasm (not
chromosomes) and have no nucleus. At
least one genetic comparison shows
Eubacteria and Archaea evolving now.
After the full genomes of all living
species are known, and understood we
will have more certainty about the
history of evolution. Many genetic
trees are based on DNA genes (sequences
of DNA that define nucleic acids or
proteins). In particular the genes for
ribosomal RNA are thought to be very
conserved over time, although perhaps
genes for reproduction, or cytoplasm,
for example may later prove to be more
conserved over time.
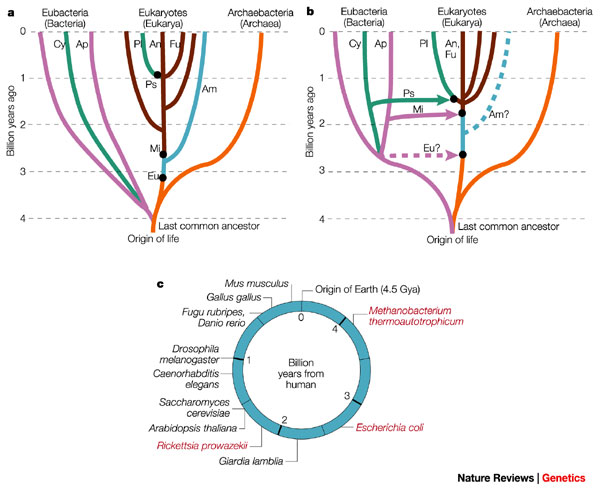
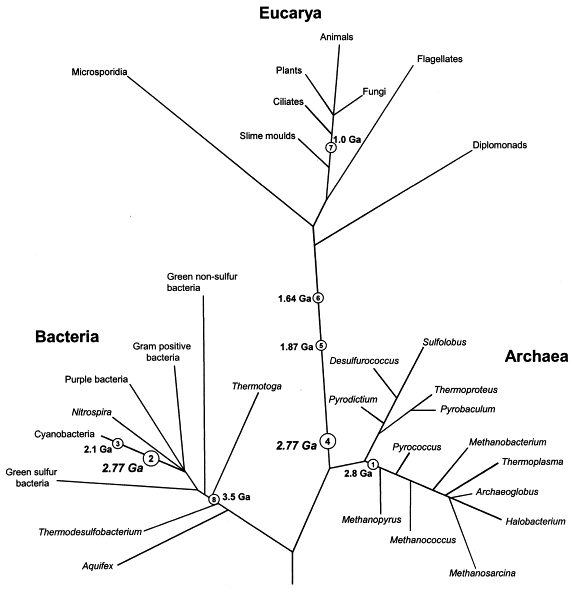
[1] Figure 1) Changing views of the
tree and timescale of life. a) An
early-1990s view, with the tree
determined mostly from ribosomal RNA
(rRNA) sequence analysis. This tree
emphasizes vertical (as opposed to
horizontal) evolution and the close
relationship between eukaryotes and the
Archaebacteria. The deep branching
(>3.5 Giga (109) years ago, Gya) of
CYANOBACTERIA (Cy) and other Eubacteria
(purple), the shallow branching
(approx1 Gya) of plants (Pl), animals
(An) and fungi (Fu), and the early
origin of mitochondria (Mi), were based
on interpretations of the geochemical
and fossil record7, 8. Some deeply
branching amitochondriate (Am) species
were believed to have arisen before the
origin of mitochondria44. Major
symbiotic events (black dots) were
introduced to explain the origin of
eukaryotic organelles42, but were not
assumed to be associated with large
transfers of genes to the host nucleus.
They were: Eu, joining of an
archaebacterium host with a eubacterium
(presumably a SPIROCHAETE) to produce
an amitochondriate eukaryote; Mi,
joining of a eukaryote host with an
alpha-proteobacterium (Ap) symbiont,
leading to the origin of mitochondria,
and plastids (Ps), joining of a
eukaryote host with a cyanobacterium
symbiont, forming the origin of
plastids on the plant lineage and
possibly on other lineages. b) The
present view, based on extensive
genomic analysis. Eukaryotes are no
longer considered to be close relatives
of Archaebacteria, but are genomic
hybrids of Archaebacteria and
Eubacteria, owing to the transfer of
large numbers of genes from the
symbiont genome to the nucleus of the
host (indicated by coloured arrows).
Other new features, largely derived
from molecular-clock studies16, 39 (Box
1), include a relatively recent origin
of Cyanobacteria (approx2.6 Gya) and
mitochondria (approx1.8 Gya), an early
origin (approx1.5 Gya) of plants,
animals and fungi, and a close
relationship between animals and fungi.
Coloured dashed lines indicate
controversial aspects of the present
view: the existence of a
premitochondrial symbiotic event and of
living amitochondriate eukaryotes,
ancestors of which never had
mitochondria. c) The times of
divergence of selected model organisms
from humans, based on molecular clocks.
For the prokaryotes (red), because of
different possible origins through
symbiotic events, divergence times
depend on the gene of interest.
source: http://www.nature.com/nrg/journa
l/v3/n11/full/nrg929_fs.html

[2] Figure 2 A phylogeny of
prokaryotes. The relationships of
selected prokaryote model organisms
based on recent studies14-19. Times of
divergence (million years ago (Mya)
plusminus one standard error) are
indicated at nodes in the tree16, 39.
Branch lengths are not proportional to
time. Phyla and phylum-level groupings
are indicated on the right.
source: http://www.nature.com/nrg/journa
l/v3/n11/full/nrg929_fs.html
evolve.

[1] tree of archaebacteria (archaea)
COPYRIGHTED
source: http://www.uni-giessen.de/~gf126
5/GROUPS/KLUG/Stammbaum.html
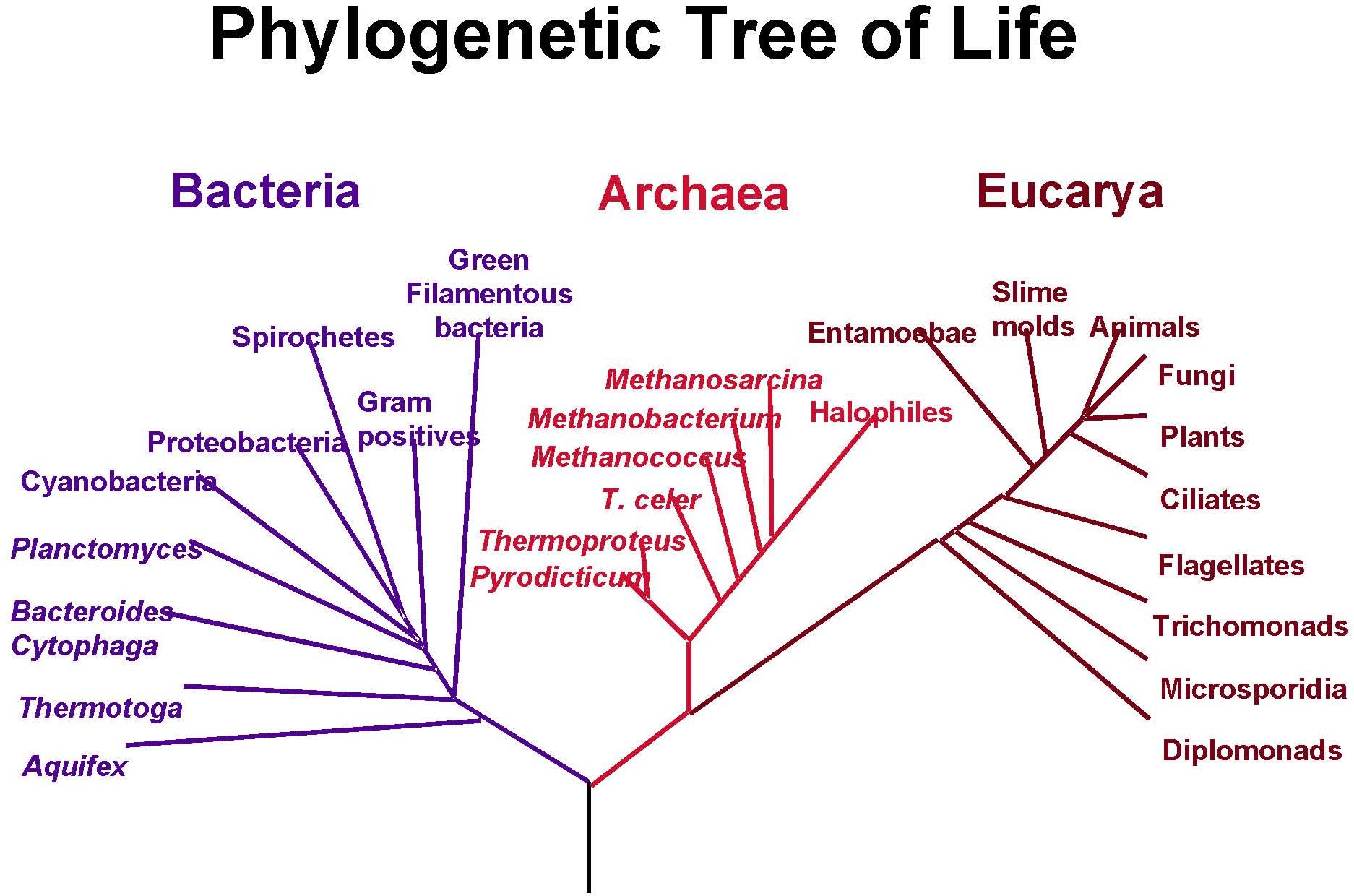
[2] A phylogenetic tree of living
things, based on RNA data, showing the
separation of bacteria, archaea, and
eukaryotes. Trees constructed with
other genes are generally similar,
although they may place some
early-branching groups very
differently, thanks to long branch
attraction. The exact relationships of
the three domains are still being
debated, as is the position of the root
of the tree. It has also been suggested
that due to lateral gene transfer, a
tree may not be the best representation
of the genetic relationships of all
organisms. NASA
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:PhylogeneticTree.jpg
evolves.
[1] tree of archaea ?
source: http://www.uni-giessen.de/~gf126
5/GROUPS/KLUG/Stammbaum.html

[2] Microscopia elettronica a
scansione dell'archeobatterio
termoacidofilo Sulfolobus solfataricus
COPYRIGHT ITALY
source: http://www.area.fi.cnr.it/r&f/n6
/ingrand.htm
(Aquifex, Thermotoga, etc.) evolve now.
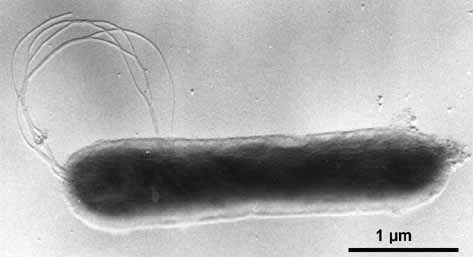
[1] Aquifex pyrophilus (platinum
shadowed). © K.O. Stetter & Reinhard
Rachel, University of Regensburg.
source: http://biology.kenyon.edu/Microb
ial_Biorealm/bacteria/aquifex/aquifex.ht
m

[2] Aquifex aeolicus. © K.O. Stetter
& Reinhard Rachel, University of
Regensburg.
source: http://biology.kenyon.edu/Microb
ial_Biorealm/bacteria/aquifex/aquifex.ht
m
also the oldest Banded Iron Formation,
on Akilia Island in Western Greenland.
The oldest evidence for life on earth
was found in this rock by measuring the
ratio of carbon 12 to carbon 13 in
grains of apatite (calcium phosphate)
from this rock. Life uses the lighter
Carbon-12 isotope and not Carbon-13 and
so the ratio of carbon-12 to carbon-13
is different from a nonliving source
(calcium carbonate or limestone).

source: nature 11/7/96
Banded Iron Formation Rocks. These
rocks are sedimentary. They are made
of iron rich chert (silicates, like
SiO2). These rocks have alternative
bands of orange or yellow and black.
In the red parts the iron is oxydized
(contains iron oxides, either hematite
{Fe2O3 = rust} or magnetite {Fe3O4]}).
These bands may have formed because
photosynthetic bacteria (in
stromatolites found in shallow ocean
shores, and purple bacteria floating in
water) produce oxygen from CO2 during
photosynthesis. When the level of
oxygen in the water became too high,
many bacteria died, and this cycle
created the BIF. But BIF also may form
naturally when photons in uv
frequencies split H2O into H2 and O2.
So perhaps the BIF bands represent
cycles of more or less uv light
reaching the earth. Perhaps the
alternating phenomenon is similar to
eukaryotic algal blooms. In any event,
this free oxygen bonded with the many
tons of iron dissolved in the water to
form insoluable iron oxide which then
fell to the ocean floor to form the
orange layers of Banded Iron Formation.
How these alternating bands are made
is not clear and has not yet been
duplicated in a lab.
This cycle of alternating orange and
black bands will continue for 2 billion
years until 1,800 million years before
now. This is the beginning of oxygen
production on earth, the atmosphere of
earth still has only small amounts of
oxygen at this time.
source: nature 11/7/96
Greenland Banded Iron Formation
sediment are evidence of the existence
of Archaea.
and Hydrocarbon molecules (alkanes)
detected in 3760 billion year old Isua
Banded Iron Formation, indicate the
possibility of photosynthetic sulfate
reducing bacteria (Archaea, for example
Sulpholobus) and Cyanobacteria living
at that time.
in Isua, Greenland Banded Iron
Formation evidence of prokaryote Oxygen
photosynthesis.
yet found. Stromatolites made by
photosynthetic bacteria found in both
Warrawoona, Western Australia, and Fig
Tree Group, South Africa.

[1] image on left is from swaziland
source: nature feb 6

source: 1986
thought to be cyanobacteria, found in
3,500 Million Year old chert from South
Africa and 3,465 Million year old Apex
chert of north-western Australia.
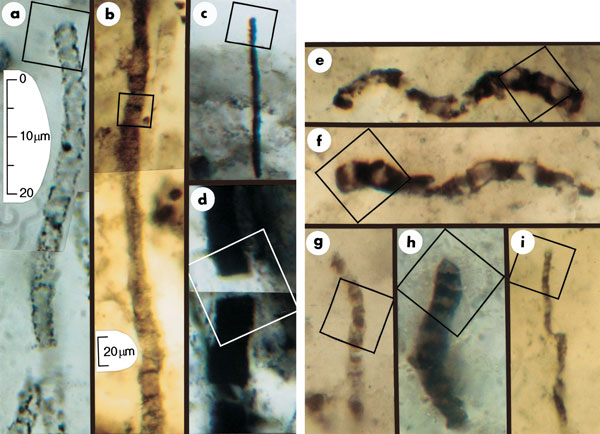
[1] Figure 1 Optical photomicrographs
showing carbonaceous (kerogenous)
filamentous microbial fossils in
petrographic thin sections of
Precambrian cherts. Scale in a
represents images in a and c-i; scale
in b represents image in b. All parts
show photomontages, which is
necessitated by the three-dimensional
preservation of the cylindrical sinuous
permineralized microbes. Squares in
each part indicate the areas for which
chemical data are presented in Figs 2
and 3. a, An unnamed cylindrical
prokaryotic filament, probably the
degraded cellular trichome or tubular
sheath of an oscillatoriacean
cyanobacterium, from the 770-Myr
Skillogalee Dolomite of South
Australia12. b, Gunflintia grandis, a
cellular probably oscillatoriacean
trichome, from the 2,100-Myr Gunflint
Formation of Ontario, Canada13. c, d,
Unnamed highly carbonized filamentous
prokaryotes from the 3,375-Myr Kromberg
Formation of South Africa14: the poorly
preserved cylindrical trichome of a
noncyanobacterial or oscillatoriacean
prokaryote (c); the disrupted,
originally cellular trichomic remnants
possibly of an Oscillatoria- or
Lyngbya-like cyanobacterium (d). e-i,
Cellular microbial filaments from the
3,465-Myr Apex chert of northwestern
Western Australia: Primaevifilum
amoenum4,5, from the collections of The
Natural History Museum (TNHM), London,
specimen V.63164[6] (e); P. amoenum4
(f); the holotype of P.
delicatulum4,5,15, TNHM V.63165[2] (g);
P. conicoterminatum5, TNHM V63164[9]
(h); the holotype of Eoleptonema apex5,
TNHM V.63729[1] (i).
source: Nature416
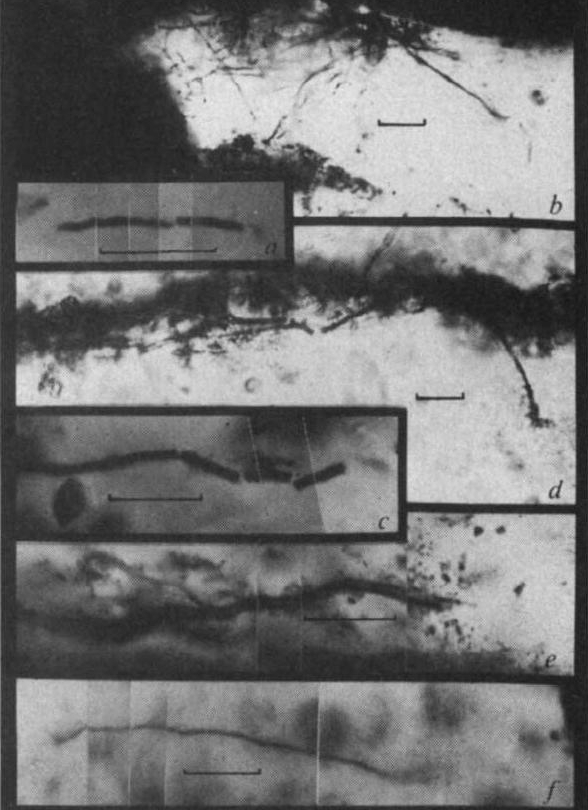
[2] Fig. 3 Filamentous microfossils:
a, cylindrical microfossil from
Hooggenoeg sample; b, threadlike and
tubular filaments extending between
laminae, Kromberg sample; c,d,e,
tubular filamnets oriented subparallel
to bedding, Kromberg sample; f,
threadlike filament flattened parallel
to bedding, Kromberg sample.
source: 73 - 76 (07 Mar 2002) Letters
to Nature
http://www.nature.com/nature/journal/v41
6/n6876/fig_tab/416073a_F1.html
evidence of moderate thermophile
sulphur reducing prokaryotes from North
Pole, Australia.
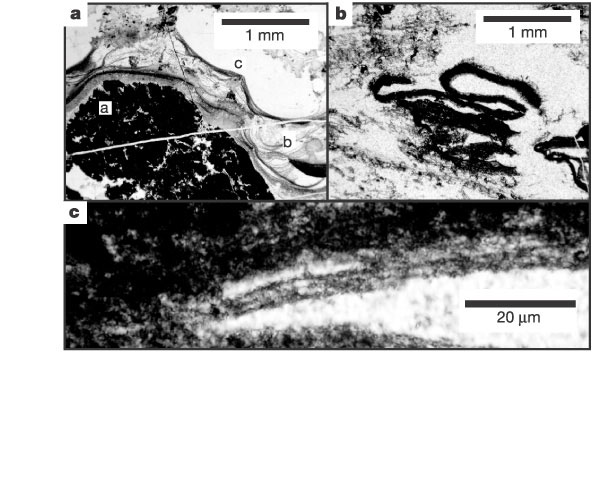
[1] get larger image
source: file:///root/web/fossils_biomark
er_science_v67_i22_nov_15_2003.html#bib9
9
bacteria.

[1] The tree is modified from ref. 2,
and abstracted from phylogenetic trees
presented in refs 26 and 27. The time
calibration points are from ref. 30,
with our additional constraint of 3.47
Gyr placed in the Bacterial domain.
Lineages housing sulphate-reducers
metabolizing at temperatures > 70 °C
are shown by broken black lines, while
lineages supporting sulphate-reducers
metabolizing at < 70 °C are shown by heavy black lines.
source: http://www.nature.com/nature/jou
rnal/v410/n6824/fig_tab/410077a0_F4.html
photosynthetic, probably anoxygenic,
bacteria that lived in mats in the
ocean date to this time.
[1] a, Dark carbonaceous laminations
draping an underlying coarse detrital
carbonaceous grain (a), showing
internal anastomosing and draping
character (b) and, at the top (c)
draping irregularities in underlying
carbonaceous laminations. b, Dark
carbonaceous laminations that have been
eroded and rolled up by currents. c,
Bundled filaments in the rolled
laminations in b [tp: they should
have clearly indicated that they are
saying that these filaments are
bacteria].
source: http://www.nature.com/nature/jou
rnal/v431/n7008/fig_tab/nature02888_F4.h
tml
Different from binary division, where a
cell is split in half, in budding, a
new complete cell is made in the
original cell, and the new cell bursts
through the cell wall, the original
cell wall must then be repaired.

[1] Evolutionary relationships of model
organisms and bacteria that show
unusual reproductive strategies. This
phylogenetic tree (a) illustrates the
diversity of organisms that use the
alternative reproductive strategies
shown in (b). Bold type indicates
complete or ongoing genome projects.
Intracellular offspring are produced by
several low-GC Gram-positive bacteria
such as Metabacterium polyspora,
Epulopiscium spp. and the segmented
filamentous bacteria (SFB). Budding and
multiple fission are found in the
proteobacterial genera Hyphomonas and
Bdellovibrio, respectively. In the case
of the Cyanobacteria, Stanieria
produces baeocytes and Chamaesiphon
produces offspring by budding.
Actinoplanes produce dispersible
offspring by multiple fission of
filaments within the sporangium.
source: http://www.nature.com/nrmicro/jo
urnal/v3/n3/full/nrmicro1096_fs.html
(Nature Reviews Microbiology 3
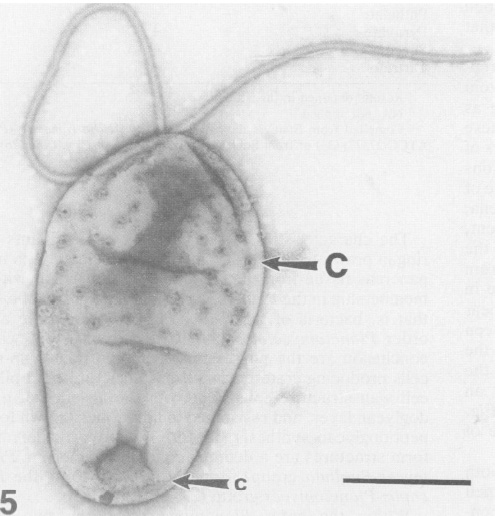
[2] Electron micrograph of a
Pirellula bacterium from giant tiger
prawn tissue (Penaeus monodon). Notice
the large crateriform structures (C) on
the cell surface and flagella. From
Fuerst et al.
source: 214-224 (2005);
doi:10.1038/nrmicro1096)
South Africa are oldest evidence of
procaryotes that reproduce by budding
and not binary fission.
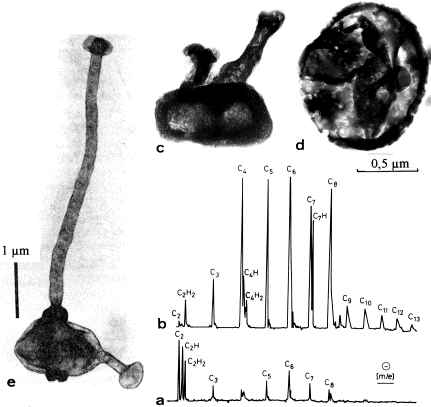
[1] Fig. 4. (a-d) Organic
microstructures from Swartkoppie chert,
South Africa (ca 3.25 Ga).
TEM-micrographs of demineralized
specimen (a,b) Laser mass spectra
(negative ions) from clusters of
similar specimens. Field of measurement
ca 1 small mu, Greekm diameter. (c,d)
TEM-micrographs from demineralized Thin
section. (e) Recent budding iron
bacterium Pedomicrobium sp. (Fig. e
from Ghiorse and Hirsch, 1979).
source: http://www.sciencedirect.com/sci
ence?_ob=MiamiCaptionURL&_method=retriev
e&_udi=B6VBP-42G6M5T-7&_image=fig6&_ba=6
&_user=4422&_coverDate=02%2F01%2F2001&_f
mt=full&_orig=browse&_cdi=5932&view=c&_a
cct=C000059600&_version=1&_urlVersion=0&
_userid=4422&md5=801178ddb930bd041063bae
7a3e0e204
found in 3235 million year old deep-sea
volcanogenic massive sulphide deposits
from the Pilbara Craton of Australia
may be oldest Archaea fossils.

[1] Photomicrographs of filaments from
the Sulphur Springs VMS deposit. Scale
bar, 10 µm. a-f, Straight, sinuous and
curved morphologies, some densely
intertwined. g, Filaments parallel to
the concentric layering. h, Filaments
oriented sub-perpendicular to
banding.
source:
G+C {Guanine and Cytosine count} Gram
positive) evolve.
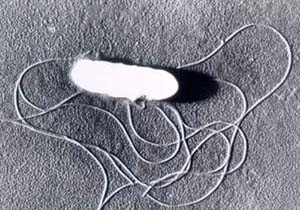
[1] Listeria monocytogenes is a
Gram-positive bacterium, in the
division Firmicutes, named for Joseph
Lister. It is motile by means of
flagella. Some studies suggest that 1
to 10% of humans may carry L.
monocytogenes in their
intestines. Researchers have found L.
monocytogenes in at least 37 mammalian
species, both domesticated and feral,
as well as in at least 17 species of
birds and possibly in some species of
fish and shellfish. Laboratories can
isolate L. monocytogenes from soil,
silage, and other environmental
sources. L. monocytogenes is quite
hardy and resists the deleterious
effects of freezing, drying, and heat
remarkably well for a bacterium that
does not form spores. Most L.
monocytogenes are pathogenic to some
degree.
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Listeria.jpg

[2] These are bacteria (about 0.3 µm
in diameter) that do not have outer
walls, only cytoplasmic membranes.
However, they do have cytoskeletal
elements that give them a distinct
non-spherical shape. They look like
schmoos that are pulled along by their
heads. How they are able to glide is a
mystery.
source: http://webmac.rowland.org/labs/b
acteria/projects_glide.html
ancestor of all Proteobacteria
(Rickettsia {mitochondria}, gonorrhoea,
Salmonella, E coli) evolving now.
[1] Figure 1. Transmission electron
micrograph of the ELB agent in XTC-2
cells. The rickettsia are free in the
cytoplasm and surrounded by an electron
transparent halo. Original
magnification X 30,000. CDC PD
source: www.cdc.gov/ncidod/
eid/vol7no1/raoultG1.htm
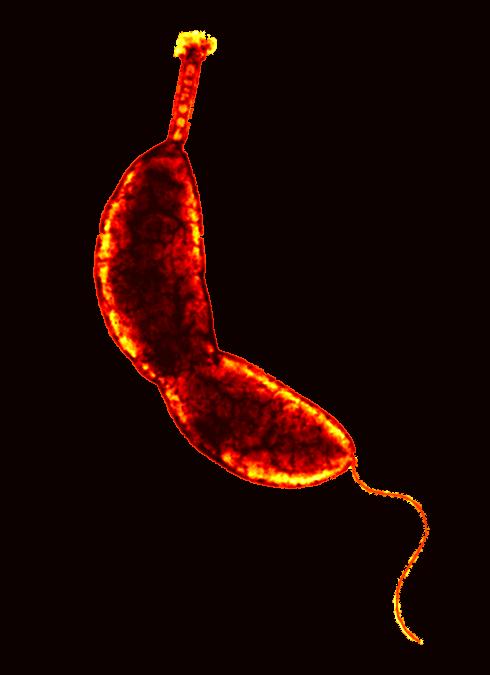
[2] Caulobacter crescentus. From
http://sunflower.bio.indiana.edu/~ybrun/
L305.html COPYRIGHTED EDU was in wiki
but appears to be removed
source: http://upload.wikimedia.org/wiki
{kind=link}
pedia/en/4/42/Caulobacter.jpg
Eubacteria Phylum, Planctomycetes
(Planctobacteria) evolving now.

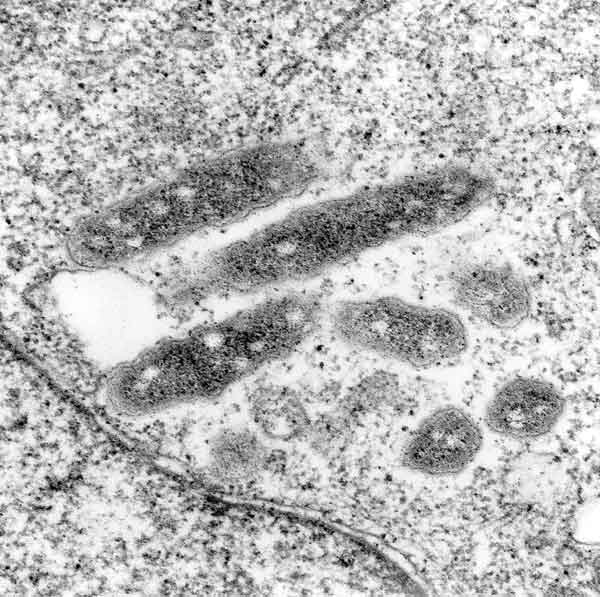
[1] Electron micrographs of cells of
new Gemmata-like and Isosphaera-like
isolates. (A) Negatively stained cell
of the Gemmata-like strain JW11-2f5
showing crateriform structures
(arrowhead) and coccoid cell
morphology. Bar marker, 200 nm. (B)
Negatively stained budding cell of
Isosphaera-like strain CJuql1 showing
uniform crateriform structures
(arrowhead) on the mother cell and
coccoid cell morphology. Bar marker,
200 nm. (C) Thin section of
Gemmata-like cryosubstituted cell of
strain JW3-8s0 showing the
double-membrane-bounded nuclear body
(NB) and nucleoid (N) enclosed within
it. Bar marker, 200 nm. (D) Thin
section of Isosphaera-like strain C2-3
possessing a fibrillar nucleoid (N)
within a cytoplasmic compartment
bounded by a single membrane (M) only.
Bar marker, 200 nm. Appl Environ
Microbiol. 2002 January; 68(1):
417-422. doi:
10.1128/AEM.68.1.417-422.2002.
source: http://www.pubmedcentral.gov/art
iclerender.fcgi?tool=pubmed&pubmedid=117
72655
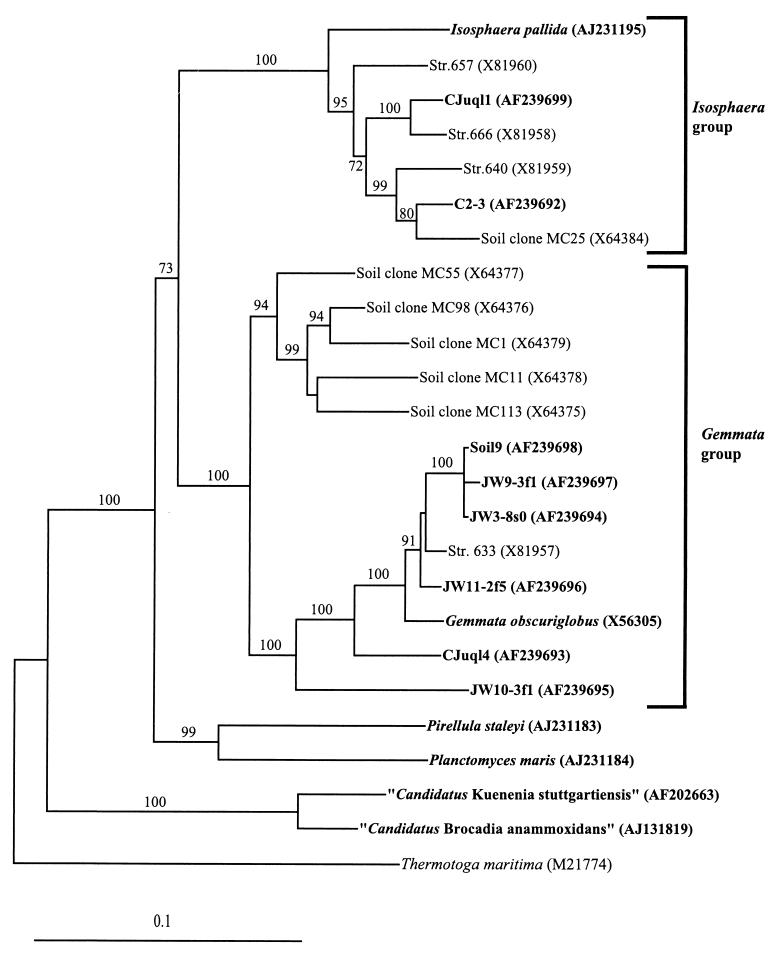
[2] Evolutionary distance tree
derived from comparative analysis of
16S rDNAs from freshwater and soil
isolates and reference strains of the
order Planctomycetales. Database
accession numbers are shown in
parentheses after species, strain, or
clone names. Bootstrap values of
greater than 70% from 100 bootstrap
resamplings from the distance analysis
are presented at nodes. Thermotoga
maritima was used as an outgroup.
Isolates from this study and
representative named species of the
planctomycetes are indicated in bold.
The scale bar represents 0.1 nucleotide
substitution per nucleotide
position. Appl Environ Microbiol.
2002 January; 68(1): 417-422. doi:
10.1128/AEM.68.1.417-422.2002.
source: http://florey.biosci.uq.edu.au/m
{kind=link}
ypa/images/fuerst2.gif
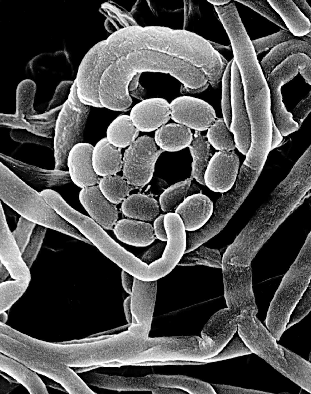
Eubacteria Phylum, Actinobacteria (high
G+C, Gram positive) evolving now.
[1] Frankia is a genus of
nitrogen-fixing soil bacteria, which
possesses a set of features that are
unique amongst symbiotic
nitrogen-fixing microorganisms,
including rhizobia, making it an
attractive taxon to study. These
heterotrophic Gram-positive bacteria
which are able to induce symbiotic
nitrogen-fixing root nodules
(actinorhizas) in a wide range of
dicotyledonous species (actinorhizal
plants), have also the capacity to fix
atmospheric nitrogen in culture and
under aerobic conditions.
source: http://www.ibmc.up.pt/webpagesgr
upos/cam/Frankia.htm
[2] Aerial mycelium and spore of
Streptomyces coelicolor. The mycelium
and the oval spores are about 1µm
wide, typical for bacteria and much
smaller than fungal hyphae and spores.
(Scanning electron micrograph, Mark
Buttner, Kim Findlay, John Innes
Centre). COPYRIGHT UK
source: http://www.sanger.ac.uk/Projects
/S_coelicolor/micro_image4.shtml
Eubacteria Phylum, Spirochaetes
(Syphilis, Lyme disease) evolving now.
[1] Syphilis is a complex, sexually
transmitted disease (STD) with a highly
variable clinical course. The disease
is caused by the bacterium, Treponema
pallidum. In the United States, 32,871
cases of syphilis, including 432 cases
of congenital syphilis, were detected
by public health officials in 2002.
Eight of the ten states with the
highest rates of syphilis are located
in the southern region of the United
States.
source: http://www.cdc.gov/nchstp/od/tus
kegee/syphilis.htm

[2] leptospirose 200x magnified with
dark-field microscope photo taken by
bluuurgh at the dutch royal tropical
institute (www.kit.nl) PD
source: http://uhavax.hartford.edu/bugl/
{kind=link}
images/Treponema%20pallidum.jpg
Eubacteria Phyla Bacteroidetes and
Chlorobi (green sulphur bacteria)
evolving now.
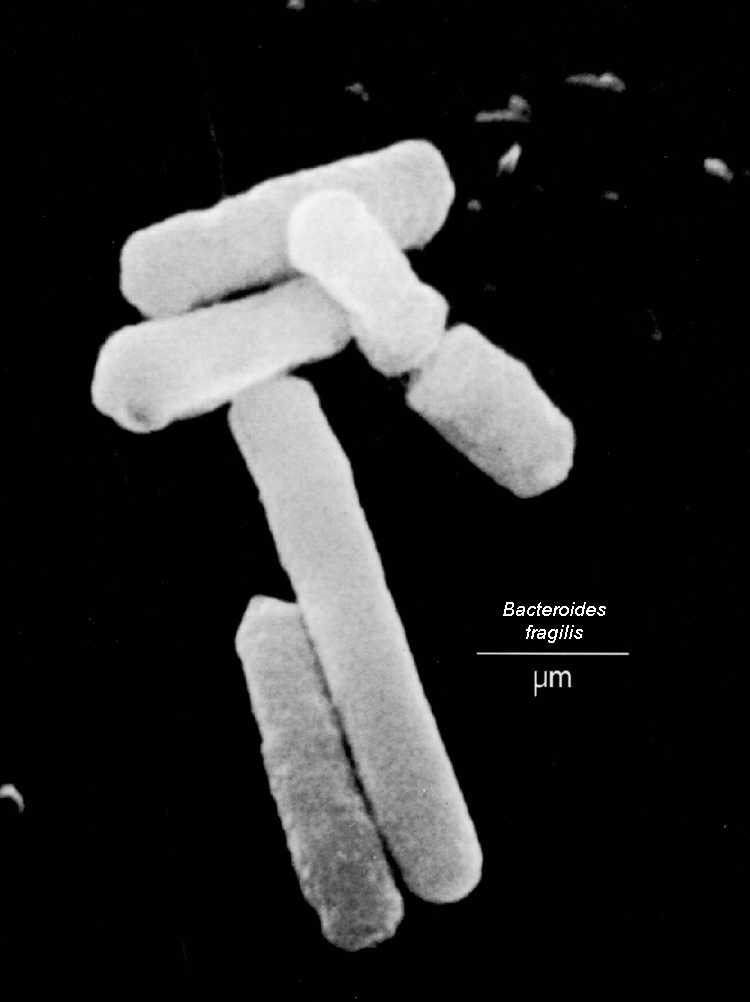
[1] Bacteroides fragilis . From the
Zdravotni University
source: http://biology.kenyon.edu/Microb
ial_Biorealm/bacteria/bacteroidete_chlor
ob_group/bacteroides/bacteroides.htm
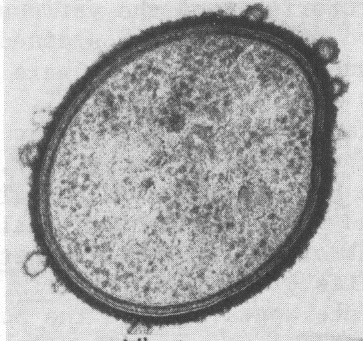
[2] Cross section of a Bacteroides
showing an outer membrane, a
peptidoglycan layer, and a cytoplasmic
membrane. From New-asthma
source: http://phil.cdc.gov/phil/details
.asp
Eubacteria Phyla Chlamydiae and
Verrucomicrobia evolving now.
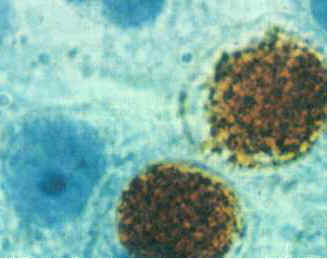
[1] Chlamydia trachomatis wiki, is
copyrighted
source: http://en.wikipedia.org/wiki/Chl
amydia_trachomatis

[2] wiki, public domain
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Chlamydophila_pneumoniae.jpg
cell membrane folds around some
molecules to form a spherical vesicle
which enters the cytoplasm, and
exocytosis, the opposite process, where
a vesicle combines with a call membrane
to empty molecules outside a cell both
evolve in an early eukaryote cell.
Eukaryote cells can now swallow
bacteria (phagocytosis) and liquid
(pinocytosis). The cells can then
(heterotrophically) use the molecules
injested (for example a bacterium) for
copying and to make ATP. This is the
first time one cell can eat a different
living cell.

[1] Pinocytosis In the process of
pinocytosis the plasma membrane froms
an invagination. What ever substance
is found within the area of
invagination is brought into the
cell. In general this material will
be dissolved in water and thus this
process is also refered to as
''cellular drinking'' to indicate that
liquids and material dissolved in
liquids are ingested by the
cell. This is opposed to the
ingestion of large particulate material
like bacteria or other cells or cell
debris.
source: http://academic.brooklyn.cuny.ed
u/biology/bio4fv/page/endocytb.htm
cytoplasm.
This cell has a nucleus, with either
single strands or a circle of DNA
inside. This is a single anaerobic
cell. This is the first protist.
This cell evolves either by:
1) two or more
bacteria joined, one with flagella
(perhaps a eubacteria) formed the
nucleus, a second formed the cytoplasm
outside the nucleus, eventually the
code to build the entire cell including
the instructions to build the symbiotic
captured bacteria was included in the
new nucleus,
2) the nucleus formed as
part of the cytoplasm lattice, perhaps
the outer wall folded in on itself
creating a double membrane, or a
membrane grew around the DNA (for
example like planctobacteria) which
provided more protection for the DNA
from the movement and digestive
activities of cytoplasm now without a
rigid cell wall,
3) a bacteria with
flagella that grew cytoplasm and a
secondary cell wall outside the
original cell wall,
4) a virus,
5) a
DNA strand from conjugation with a
different prokaryote stored in a
vesicle.
There are key features that are
different from eukaryotes and
prokaryotes:
1) Eukaryotes have a nucleus,
prokaryotes do not.
2) DNA in eukaryotes is
in the form of chromosomes, in
prokaryotes the DNA is in a circle.
3)
Eukaryotes can do endocytosis, fold
their cell membrane around some
external object and injest the object,
prokaryotes can not.
4) Eukaryotes have a
membrane lattice of proteins, actin and
myacin, prokaryotes do not.
5) Eukaryotes
have an endoplasmic reticulum and golgi
body.
6) Eukaryotes reproduce asexually by
dual binary division (both nucleus and
cell divide by binary division),
budding, or mitosis, prokaryotes
reproduce by budding or binary
division.
If the nucleus is an engulfed
prokaryote, this cell inherits the
processes of nuclear DNA duplication
and nucleus division (karyokinesis)
from prokaryote binary division.
Initially, both the nucleus and cell
divide by binary division.

[1]
http://www.regx.de/m_organisms.php#planc
to
source: http://www.regx.de/m_organisms.p
hp#plancto
[2]
http://www.mansfield.ohio-state.edu/~sab
edon/biol1080.htm
source: http://www.mansfield.ohio-state.
edu/~sabedon/biol1080.htm
single circular chromosome to linear
chromosomes.
Possibly the prokaryote ancestor of the
first eukaryote had linear chromosomes
since some prokaryotes (although very
few) are known to have linear
chromosomes instead of or in addition
to a single circular chromosome.
undulipodium) evolves on early single
cell eukaryotes.
constantly synthesizing DNA and then
having a division period (as is the
case for all known prokaryotes), but
this cell has a period in between cell
division and DNA synthesis where DNA
synthesis is not performed. Later some
cells develop a stage after synthesis
and before cell division.
a prokaryote, synchronized duplication
and division of organelle-nucleus and
cytoplasm of early eukaryote cell
evolves. Before this, eukaryote cell
division usually results in one cell
with no organelle-nuclei and a second
cell with 2 organelle-nuclei. Perhaps
the organelle-nuclei attach to the
outer cell membrane in the same way the
cytoplasmic DNA do, which allows new
cytoplasm growth to separate the two
organelle-nucleus in addition to the
cytoplasmic DNA.
source:
source:
haploid (single set of chomosomes)
eukaryote nucleus, evolves in
eukaryotes. Before mitosis, there is a
synthesis stage where DNA in the form
of chromosomes are duplicated in the
nucleus before the nucleus and cell
divide.

[1] Mitosis divides genetic information
during cell division Source:
http://www.ncbi.nlm.nih.gov/About/primer
/genetics_cell.html This image is
from the Science Primer, a work of the
National Center for Biotechnology
Information, part of the National
Institutes of Health. As a work of the
U.S. federal government, the image is
in the public domain.
source: http://en.wikipedia.org/wiki/Mit
osis
[2] Prophase: The two round objects
above the nucleus are the centrosomes.
Note the condensed chromatin. from
Gray's Anatomy. Unless stated
otherwise, it is from the online
edition of the 20th U.S. edition of
Gray's Anatomy of the Human Body,
originally published in 1918. Online
editions can be found on Bartleby and
also on Yahoo!
source:
syngamy, gametogamy) evolves in
protists. Haploid (1 set of
chromosomes) eukaryote cells merge and
then their nuclei merge (karyogamy) to
form the first diploid (2 sets of
chromosomes) cells (the first zygote).
This fusion of 2 haploid cells results
in the first diploid single-celled
organism, which then immediately
divides (both nucleus and cytoplasm by
single-division meiosis) back to two
haploid cells.
Possibly first, only cytoplasmic
merging happened with nuclear merging
(karyogamy) and nuclear division
(karyokinesis) evolving later.
Now, two cells
with different DNA can mix providing
more chance of variety/mutation. Two
chromosome sets provides a backup copy
of important genes (sequences that code
for proteins, or nucleic acids) that
might be lost with only a set of single
chromosomes.
The life cycle of future organisms will
now have two phases, a gamophase (from
n to 2n (until syngamy)), and zygophase
(from 2n to n (until meiosis)). Gamoid
cells are not haploid in polyploid
organisms.
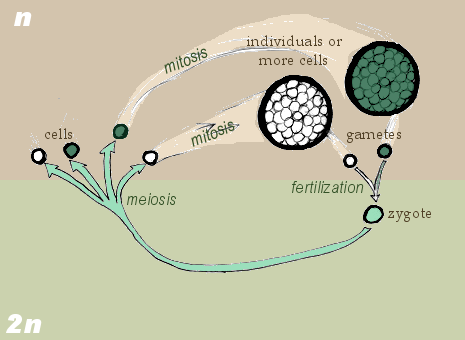
[1] Zygotic Meiosis. GNU
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Zygotic_meiosis.png

[2] Gametic Meiosis. GNU
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Gametic_meiosis.png
duplication and a cell division of a
diploid cell into 2 haploid cells)
evolves.
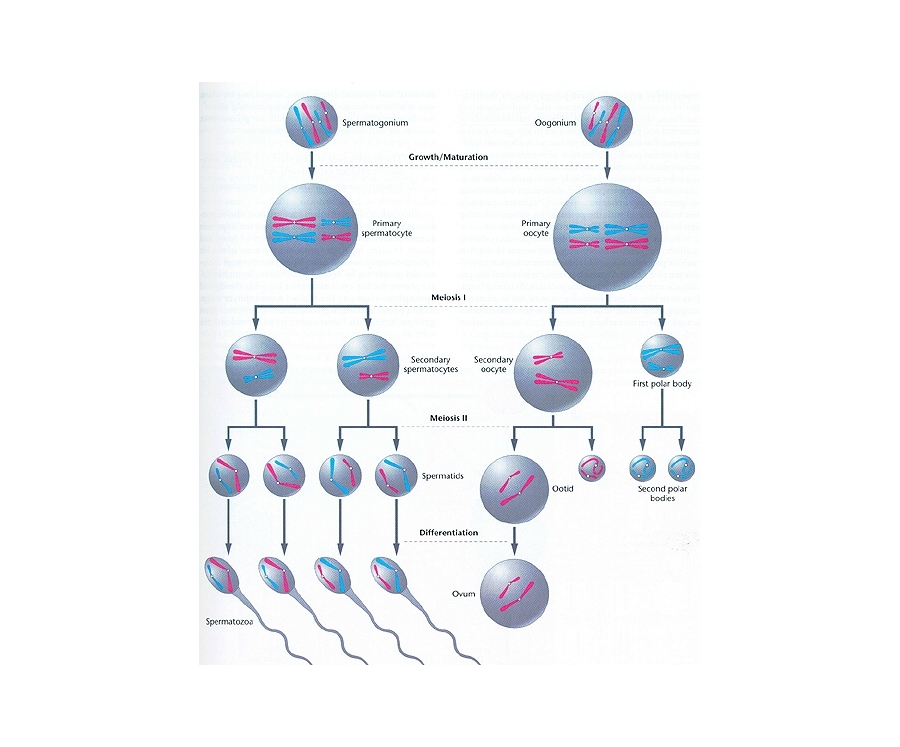
[1] GametoGenesis. COPYRIGHTED EDU
source: http://www.bio.miami.edu/dana/10
{kind=link}
4/gametogenesis.jpg

[2] Sexual cycle oxymonas, identical
to saccinobaculus, one step meiosis.
haploid. COPYRIGHTED CANADA
source: http://www.zoology.ubc.ca/~redfi
{kind=link}
eld/clevelan/oxymonas.GIF
2 haploid cells fuse) evolves.
This begins the "diplontic" life cycle
(with gametic meiosis), where diploid
cells (a zygote) can copy asexually
through mitosis after merging. This
organism, when haploid, cannot do
mitosis (presumably haploid gamete
mitosis will evolve much later in brown
algae), and this is still true in all
descendents (including humans) of this
single celled organism.
(cell and nucleus fusion) between two
isogamous (same size) gametes but which
have 2 different (+ and -) forms
(genders).
between two different size gamete cells
(heterogamy or anisogamy) evolves in
protists.
sterols, molecules made by mitochondria
in eukaryotes) found in northwestern
Australia.
evolves.
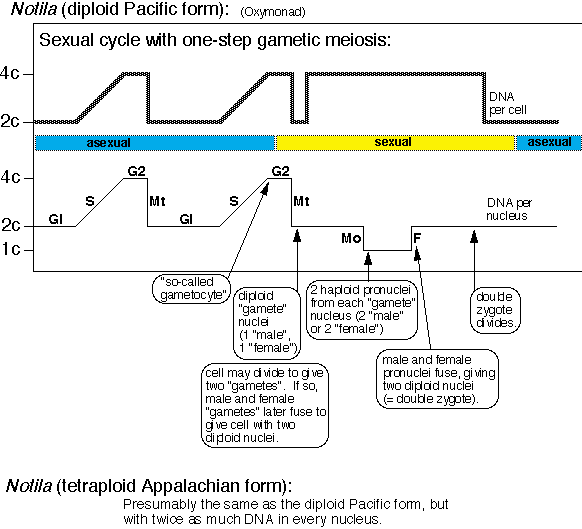
[1] The Oxymonad, Notila (diploid
Pacific form) life cycle. COPYRIGHTED
source: http://www.zoology.ubc.ca/~redfi
{kind=link}
eld/clevelan/notila.GIF
cell divisions with no stage in between
which result in one diplid cell
dividing into four haploid cells)
evolves.
[1] GametoGenesis. COPYRIGHTED EDU
source: http://www.bio.miami.edu/dana/10
4/gametogenesis.jpg
[2] Sexual cycle oxymonas, identical
to saccinobaculus, one step meiosis.
haploid. COPYRIGHTED CANADA
source: http://www.zoology.ubc.ca/~redfi
eld/clevelan/oxymonas.GIF
(Thermus Aquaticus {used in PCR},
Deinococcus radiodurans {can survive
long exposure to radiation}) evolve
now.
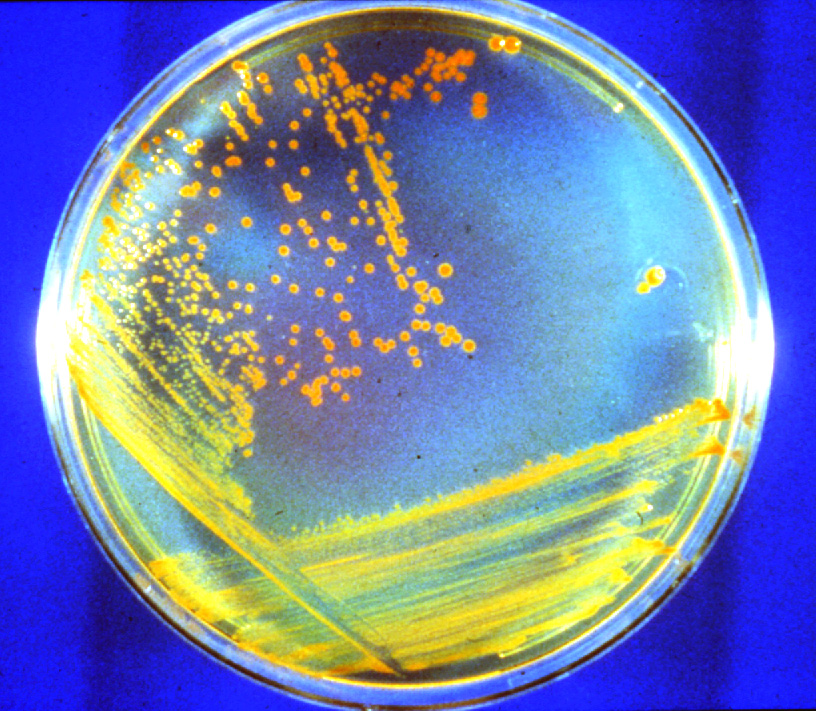
[1] D. radiodurans growing on a
nutrient agar plate. The red color is
due to carotenoid pigment. Links to
816x711-pixel, 351KB JPG. Credit: M.
Daly, Uniformed Services University of
the Health Sciences NASA
source: http://science.nasa.gov/newhome/
{kind=link}
headlines/images/conan/D_rad_dish.jpg
[2] Photomicrograph of Deinococcus
radiodurans, from
www.ornl.gov/ORNLReview/ v34 The Oak
Ridge National Laboratory United
States Federal Government This work
is in the public domain because it is a
work of the United States Federal
Government. This applies worldwide. See
Copyright.
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Deinococcus.jpg
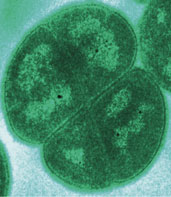
Eubacteria phylum, Cyanobacteria
(ancestor of all eukaryote chloroplasts
{plastids}) evolving now. There is a
conflict between the interpretation of
the geological and the genetic evidence
as to if oxygen photosynthesis and
cyanobacteria evolved earlier around
3800mybn or here at 2500mybn.

[1] Oscillatoria COPYRIGHTED EDU
source: http://www.stcsc.edu/ecology/alg
{kind=link}
ae/oscillatoria.jpg

[2] Lyngbya COPYRIGHTED EDU
source: http://www.stanford.edu/~bohanna
{kind=link}
n/Media/LYNGB5.jpg
Non-Sulphur) evolve now.

[1] Chloroflexus photomicrograph from
Doe Joint Genome Institute of US Dept
Energy PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Chlorofl.jpg
Era.
appear in many places.
million years starts now.
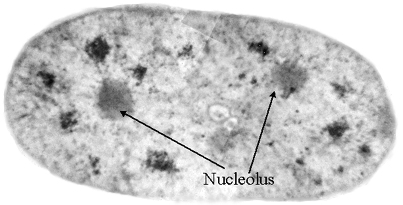
nucleus that makes ribosomes, evolves.
[1] Nucleolus, COPYRIGHTED
source: http://www.eccentrix.com/members
{kind=link}
/chempics/Slike/cell/Nucleolus.jpg
[2] With the combination of x-rays
from the Advanced Light Source and a
new protein-labeling technique,
scientists can see the distribution of
the nucleoli within the nucleus of a
mammary epithelial cell. USG PD
source: http://www.lbl.gov/Science-Artic
les/Archive/xray-inside-cells.html
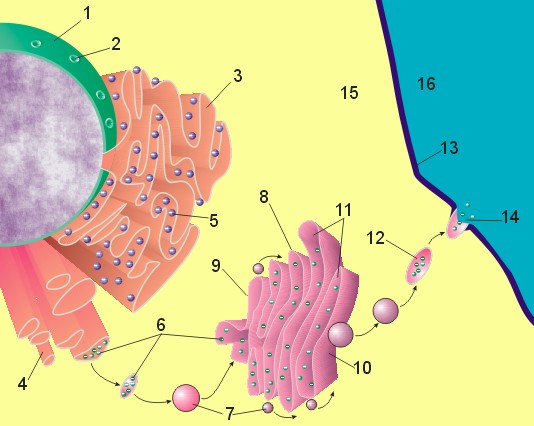
reticulum evolves in eukaryote cell.

[1] Figure 1 : Image of nucleus,
endoplasmic reticulum and Golgi
apparatus. (1) Nucleus. (2) Nuclear
pore. (3) Rough endoplasmic reticulum
(RER). (4) Smooth endoplasmic reticulum
(SER). (5) Ribosome on the rough ER.
(6) Proteins that are transported. (7)
Transport vesicle. (8) Golgi apparatus.
(9) Cis face of the Golgi apparatus.
(10) Trans face of the Golgi apparatus.
(11) Cisternae of the Golgi
apparatus. I am the copyright holder
of that image (I might even have the
CorelDraw file around somewhere:-), and
I hereby place the image and all
partial images created from it in the
public domain. So, you are free to use
it any way you like. In fact, I am
delighted that one of my drawings makes
it into print! I can mail you the
.cdr file, if you like (and if I can
find it), if you need a better
resolution for printing. Yours, Magnus
Manske Source: [1]. See also
User:Magnus Manske
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Nucleus_ER_golgi.jpg
dictyosome) evolves in eukaryote cell.
[1] Figure 1: Image of nucleus,
endoplasmic reticulum and Golgi
apparatus: (1) Nucleus, (2) Nuclear
pore, (3) Rough endoplasmic reticulum
(RER), (4) Smooth endoplasmic reticulum
(SER), (5) Ribosome on the rough ER,
(6) Proteins that are transported, (7)
Transport vesicle, (8) Golgi apparatus,
(9) Cis face of the Golgi apparatus,
(10) Trans face of the Golgi apparatus,
(11) Cisternae of the Golgi apparatus,
(12) Secretory vesicle, (13) Plasma
membrane, (14) Exocytosis, (15)
Cytoplasm, (16) Extracellular space.
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Nucleus_ER_golgi_ex.jpg
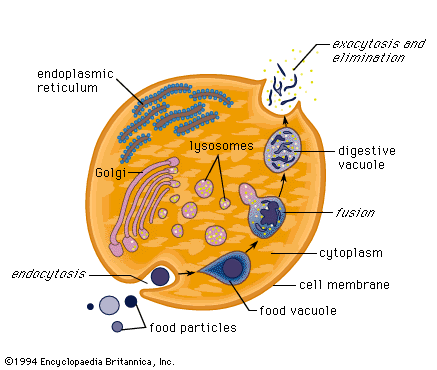
makes lysosomes which fuse with
endosomes. The various molecules in
lysosomes digest the contents of the
endosome, which then exits the cell as
waste.
[1] Figure 1: Image of nucleus,
endoplasmic reticulum and Golgi
apparatus: (1) Nucleus, (2) Nuclear
pore, (3) Rough endoplasmic reticulum
(RER), (4) Smooth endoplasmic reticulum
(SER), (5) Ribosome on the rough ER,
(6) Proteins that are transported, (7)
Transport vesicle, (8) Golgi apparatus,
(9) Cis face of the Golgi apparatus,
(10) Trans face of the Golgi apparatus,
(11) Cisternae of the Golgi apparatus,
(12) Secretory vesicle, (13) Plasma
membrane, (14) Exocytosis, (15)
Cytoplasm, (16) Extracellular space.
source: http://sun.menloschool.org/~cwea
ver/cells/e/lysosomes/
source: http://en.wikipedia.org/wiki/Ima
ge:Nucleus_ER_golgi_ex.jpg
that can only live in other bacteria,
closely related to Rickettsia
prowazekii, an aerobic
alpha-proteobacteria that causes
louse-borne typhus, enters an early
eukaryote cell. As time continues a
symbiotic relationship evolves, where
the Rickettsia forms the mitochondria,
organelles of every euokaryote cell.
The mitochondria perform the Acid
Citric Cycle (Krebs Cycle), using
oxygen to breakdown glucose into CO2
and H2O, and provide up 38 ATP
molecules. Mitochondria reproduce by
themselves, and are not created by the
DNA in the cell nucleus. As time
continues some of the DNA from the
mitochondria merges with the cell
nucleus DNA. Mitochondria produce
sterol used to make the eukaryote cell
wall flexible. Because mitochondria
need Oxygen, but the level of oxygen is
very low on earth, oxygen may be
provided by photosynthesizing
cyanobacteria living near these cells.
All eukaryotes alive today either have
mitochondria except the amitochondriate
excavates (metamonads), the most
ancient of the eukaryotes alive today.
That parabasalids have hydrogenosomes,
anaerobic organelles that seem to have
evolved from mitochondria, many people
think amitochondriate species lost
their mitochondria over time.

[1] Phylogenetic hypothesis of the
eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas.
source: http://nar.oxfordjournals.org/co
{kind=link}
ntent/vol26/issue4/images/gkb18201.gif

[2] Figure 1 Phylogenetic tree of
eukaryotes based on ultrastructural and
molecular data. Organisms are
sub-divided into main groups as
discussed in the text. Only a few
representative species for which
complete (or almost complete) mtDNA
sequences are known are shown in each
lineage. In some cases, line drawings
or actual pictures of the organisms are
provided (Acanthamoeba, M. Nagata; URL:
http://protist.i.hosei.ac.jp/PDB/PCD3379
/htmls/21.html; Allomyces, Tom Volk;
URL:
http://botit.botany.wisc.edu/images/332/
Chytridiomycota/Allomyces_r_So_pa/A._arb
uscula_pit._sporangia_tjv.html;
Amoebidium, URL:
http://cgdc3.igmors.upsud.fr/microbiolog
ie/mesomycetozoaires.htm; Marchantia,
URL:
http://www.science.siu.edu/landplants/He
patophyta/images/March.female.JPEG
Scenedesmus, Entwisle et al.,
http://www.rbgsyd.gov.au/_data/page/1824
/Scenedesmus.gif). The color-coding of
the main groups (alternating between
dark and light blue) on the outer
circle corresponds to the color-coding
of the species names. Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
molecular data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional sequence data. [t:
why not color code or add which type of
mito?]
source: http://arjournals.annualreviews.
org/doi/full/10.1146/annurev.genet.37.11
0801.142526
Unikonts (one cilium). Bikonts (also
called anterokonts for having anterior
{forward facing} cilia) will evolve
into the vast majority of the Protist
and all of the Plant Kingdoms. The
Unikonts will evolve into the ameobozoa
(tenatively), and the opisthokonts
(ancestrally posterior cilium) which
include the entire Fungi and Animal
Kingdoms.
[1] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas.
source:
a mineral that cannot exist for much
time if exposed to oxygen, indicating
that free oxygen is accumulating in the
air of earth for the first time.
on land, begin here and are evidence of
more free oxygen in the air of earth.

[1]
http://www.kgs.ukans.edu/Extension/redhi
lls/redhills.html
source:
oldest line of eukaryotes still in
existence, the oldest living protists,
in the Phylum "Metamonada" (Excavates)
originating now. This is where the
eukaryote line is created and separates
from the archaebacteria (archaea) line.
Most of these species have an
excavated ventral feeding groove, and
all lack mitochondria. Mitochondria
are thought to have been lost
secondarily, although this is not
certain.
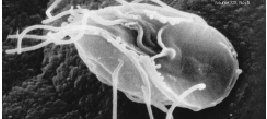
[1] Giardia lamblia, a parasitic
flagellate that causes giardiasis.
Image from public domain source at
http://www.nigms.nih.gov/news/releases/i
mages/para.jpg
source: http://www.nigms.nih.gov/news/re
{kind=link}
leases/images/para.jpg
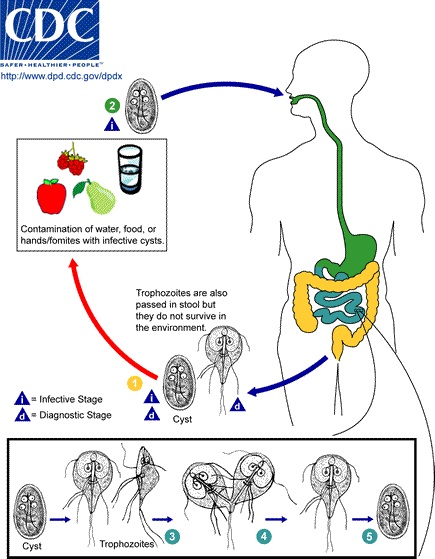
[2] . The cysts are hardy and can
survive several months in cold water.
Infection occurs by the ingestion of
cysts in contaminated water, food, or
by the fecal-oral route (hands or
fomites) . In the small intestine,
excystation releases trophozoites (each
cyst produces two trophozoites) .
Trophozoites multiply by longitudinal
binary fission, remaining in the lumen
of the proximal small bowel where they
can be free or attached to the mucosa
by a ventral sucking disk .
Encystation occurs as the parasites
transit toward the colon. The cyst is
the stage found most commonly in
nondiarrheal feces . Because the cysts
are infectious when passed in the stool
or shortly afterward, person-to-person
transmission is possible. While
animals are infected with Giardia,
their importance as a reservoir is
unclear.
source: http://www.dpd.cdc.gov/dpdx/HTML
/Giardiasis.asp?body=Frames/G-L/Giardias
is/body_Giardiasis_page1.htm
Eukaryote Phylum "Loukozoa" (Jakobea
and Malawimonadea) originating now.
These species have mitochondria with
tubular cristae, and are the most
ancient species that still have
mitochondria.
This species is the most ancient known
species to have a shell. This first
hard shells (lorika) made of calcium
carbonate (Calcite CaCO3), plates of
silica (SiO2), or carbon-based
molecules evolve around the
single-celled species living in the
ocean.
Perhaps this shell served to protect
the cell from external damage from
being eaten by other eukaryotes
(zooplankton), infection by bacteria or
viruses, control of buoyancy, to filter
UV light, against damage by non-living
sources.

[1] Histiona. This drawing was made by
D. J. Patterson. COPYRIGHTED EDU
source: http://microscope.mbl.edu/script
s/microscope.php?func=imgDetail&imageID=
3479
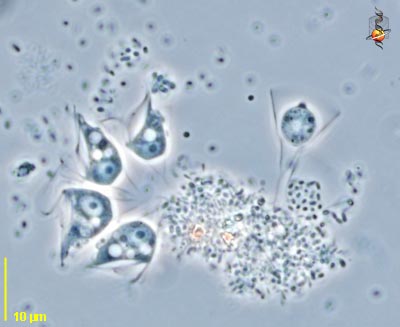
[2] Histiona (hist-ee-own-a) is a
jakobid flagellate related to Jakoba.
As with other excavates, there is a
ventral groove and the flagella insert
at the head of the groove. There are
two flagella, one lying in the groove
and one curving outwards from the point
of insertion. The margins of the groove
can be mistaken for flagella. Unlike
most other excavates, Histiona sits in
a stalked lorica when feeding. Lorica
with a cyst is evident. Phase contrast.
This picture was taken by David
Patterson, Linda Amaral Zettler, Mike
Peglar and Tom Nerad from cultures and
other materials maintained at the
American Type Culture Collection during
2001. COPYRIGHTED EDU
source: http://microscope.mbl.edu/script
s/microscope.php?func=imgDetail&imageID=
435
mitochondria split from the tubular
christae line.
This is the origin of the
Discicristata: species that have
discoid mitochondrial cristae and, in
some cases, a deep (excavated) ventral
feeding groove.
[1] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas.
source: http://nar.oxfordjournals.org/co
ntent/vol26/issue4/images/gkb18201.gif
Diplobiontic) life cycle (organism with
both diploid and haploid "alternate
life stages" that reproduce asexually
by mitosis) with "sporic meiosis"
evolves.
In this life cycle haploid gametes fuse
to form a diploid zygote which divides
by meiosis producing haploid spores
that produce (differentiate?) gametes,
starting the cycle again.
Initially these species are single
celled in both stages (like
Haptophyta).
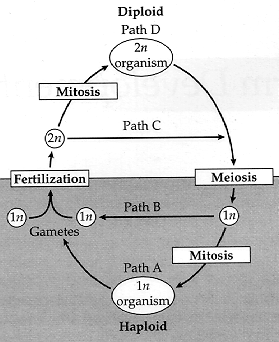
[1] Figure 23.1.Plants have
haplodiplontic life cycles that involve
mitotic divisions (resulting in
multicellularity) in both the haploid
and diploid generations (paths A and
D). Most animals are diplontic and
undergo mitosis only in the diploid
generation (paths B and D).
Multicellular organisms with haplontic
life cycles follow paths A and C.
COPYRIGHTED EDU
source: http://zygote.swarthmore.edu/pla
{kind=link}
ntfig1.gif
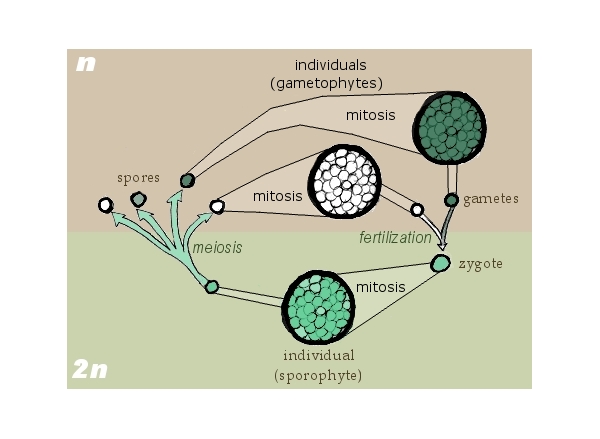
[2] Drawn by self for Biological life
cycle Based on Freeman & Worth's
Biology of Plants (p. 171). GNU
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Sporic_meiosis.png
with flat christae evolve from those
with tubular christae.
[1] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas.
source: http://nar.oxfordjournals.org/co
ntent/vol26/issue4/images/gkb18201.gif
primitive living members of the Phylum
"Euglenozoa" (euglenids, leishmania,
trypanosomes, kinetoplastids) evolved
at this time.
This is the oldest eukaryote to exhibit
colonialism. Perhaps eukaryote
colonialism is partially or fully
inherited from prokaryotes, but
colonialism may have evolved
independently again in eukaryotes.
This is the most ancient species known
to have a cell covering, which is of
the type "pellicle".

[1] euglena
source: http://www.fcps.k12.va.us/Stratf
ordLandingES/Ecology/mpages/euglena.htm
[2] euglena
source: http://protist.i.hosei.ac.jp/PDB
{kind=link}
/Images/Mastigophora/Euglena/genus1L.jpg
Phylum "Percolozoa" (also called
"Heterolobosea") (acrasid slime molds)
evolved at this time.
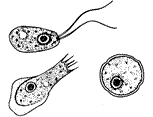
[1] Stages of Naegleria fowleri, a
member of the Percolozoa. Adapted from
Image:Free-living amebic
infections.gif, which is from the CDC.
PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Naegleria.png

[2] CLASS Heterolobosea ORDER
Schizopyrenida Heteramoeba: The
flagellated form is large (30
�m), two flagella, an elongate
cytostome curving around the anterior
of the cell and forming a groove.
Nucleus with peripheral chromatin.
Probably feeds and divides as a
flagellate. One species. This genus is
most like Paratetramitus from which it
can be distinguished by peripheral
location of chromatin material. Cysts
without pores, excystment through a
weak region of wall. Marine.
Heteramoeba (het-err-a-me-ba) a naked
heterolobose amoeba, distinguished from
other types of naked amoebae with
lobose pseudopodia largely by
ultrastructural features, but trophic
heterolobose amoebae tend to form their
pseudopodially suddenly rather than
progressively. Phase contrast. This
picture was taken by David Patterson,
Linda Amaral Zettler, Mike Peglar and
Tom Nerad from cultures and other
materials maintained at the American
Type Culture Collection during 2001.
NONCOMMERCIAL USE
source: http://microscope.mbl.edu/script
s/microscope.php?func=imgDetail&imageID=
413
protist.
Multicellularity is a very important
event in the evolution of life on
earth. With multicellular organisms,
larger sized organisms could evolve.
There are many uncertainties
surrounding the origin of
multicellularity. Multicellularity may
have evolved independently for Plants,
Fungi and Animals, or multicellularity
may have evolved only once in
eukaryotes.
The key feature of this cell is that a
multicellular organism is made from a
single cell and the multicellular
organism is not a collection of
independent cells (colonialism). The
main difference between this organism
and single-celled organisms is the way
the cells stay fastened together after
cell division.
Which species was the first
multicellular species is not clear.
Multicellularity is found in all 3 life
cycles (haplontic, diplontic,
haplodiplontic). The 3 main life cycle
types (haplontic, etc.) probably
evolved in single cell species before
multicellularity evolved. If
multicellularity evolved once and is
inherited, perhaps all multicellular
organism descended from a single
haplodiplontic organism.
These multicellular organisms have
undifferentiated cells in the
multicellular stage (all cells in the
haploid or diploid multicellular
organism are made of one kind of cell).
differentiation evolves.
Multicellular organisms are no longer
all haploid or diploid gamete producing
cells (or spore producing if
haplodiplontic), but are made of gamete
(or spore) producing cells in addition
to somatic cells which copy asexually
through mitosis.
Now, in addition to being large
multicell organisms, multicellular
organisms can have differentiated cells
that form a variety of different shaped
structures, and perform different
functions.
ancestor of the "Chromalveolates"
evolving now. Chromalveolates include
the Chromista and Alveolata. The
Chromista include the 3 Phyla
Haptophyta, Cryptophyta (Cryptomonads),
and Heterokontophyta (brown algae
{kelp}, diatoms, water molds).
Alveolata include the 3 Phyla
Dinoflagellata, Apicomplexa (Malaria,
Toxoplasmosis), and Ciliophora
(ciliates).
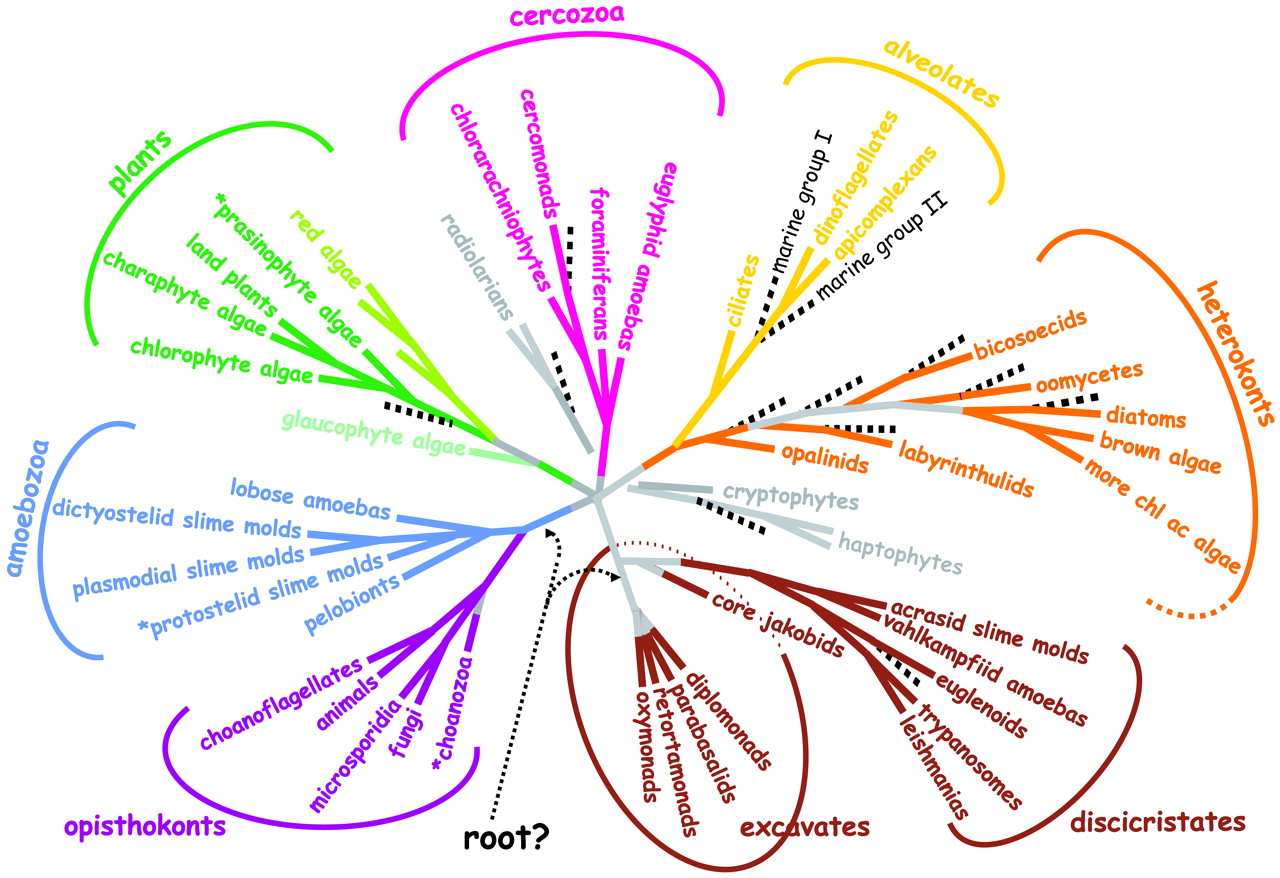
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703

[2] Beautiful marine diatoms as seen
through a microscope. These tiny
phytoplankton are encased within a
silicate cell wall. Image ID: corp2365,
NOAA Corps Collection Photographer: Dr.
Neil Sullivan, University of Southern
Calif. NOAA This image is a work of
the National Oceanic and Atmospheric
Administration, taken or made during
the course of an xxxxx? official
duties. As works of the U.S. federal
government, all NOAA images are in the
public domain.
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Diatoms_through_the_microscope.jpg
ancestor of Chromalveolate Phlyum
Haptophyta evolving now.
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Emiliania huxleyi, a
coccolithophore. Photo courtesy Dr.
Markus Geisen - photographer, and The
Natural History Museum. PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Emiliania_huxleyi_3.jpg
ancestor of the Chromalveolate Phylum
"Cryptophyta" (Cryptomonads) evolving
now.
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
ancestor of the Chromalveolate Phylum
"Heterokontophyta" (Heterokonts also
called Stramenopiles) evolving now.
Heterokonts include brown algae,
diatoms, golden algae, axodines,
yellow-green algae, water moulds and
slime nets.
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
Algae (Phaeophyta) evolves now.
Brown Algae is the most genetically
ancient multicellular organism still
living on earth. In addition to being
first to evolve multicellularity, cell
differentiation (cells of different
types) is already present in all brown
algae.
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
evolve.
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
molds (Oomycetes OemISETEZ) evolve.
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
(Ciliates, Dinoflagellates,
Apicomplexans) evolve.
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
[1] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas. COPYRIGHTED
source: http://nar.oxfordjournals.org/cg
i/content/full/26/4/865
(the Phyla "Radiolaria", "Cercozoa",
and "Foraminifera") evolve now.
This marks the beginning of the
protists described as "amoeboid",
because they have pseudopods.
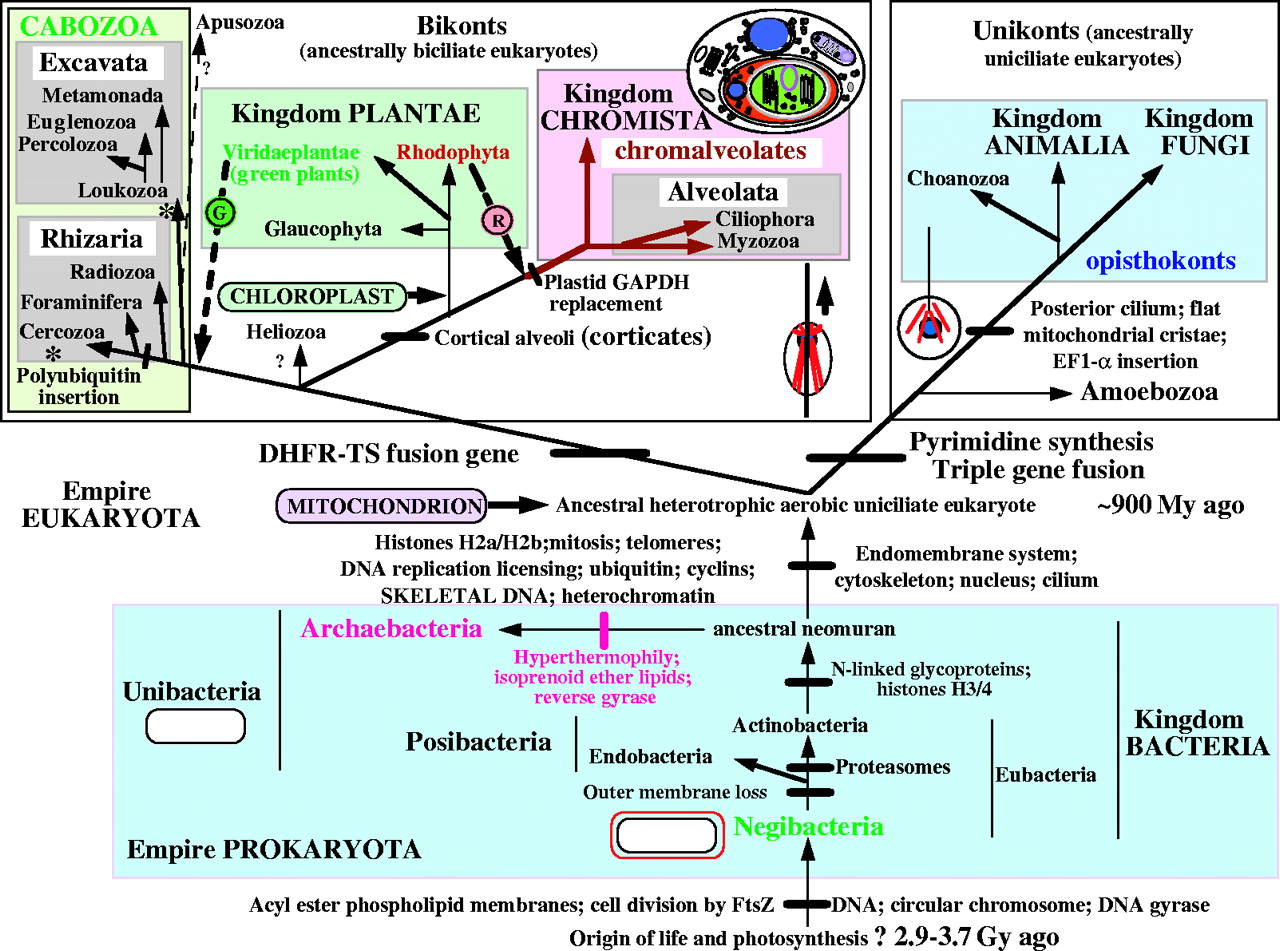
[1] FIG. 2. The tree of life based on
molecular, ultrastructural and
palaeontological evidence. Contrary to
widespread assumptions, the root is
among the eubacteria, probably within
the double-enveloped Negibacteria, not
between eubacteria and archaebacteria
(Cavalier-Smith, 2002b); it may lie
between Eobacteria and other
Negibacteria (Cavalier-Smith, 2002b).
The position of the eukaryotic root has
been nearly as controversial, but is
less hard to establish: it probably
lies between unikonts and bikonts (Lang
et al., 2002; Stechmann and
Cavalier-Smith, 2002, 2003). For
clarity the basal eukaryotic kingdom
Protozoa is not labelled; it comprises
four major groups (alveolates, cabozoa,
Amoebozoa and Choanozoa) plus the small
bikont phylum Apusozoa of unclear
precise position; whether Heliozoa are
protozoa as shown or chromists is
uncertain (Cavalier-Smith, 2003b).
Symbiogenetic cell enslavement occurred
four or five times: in the origin of
mitochondria and chloroplasts from
different negibacteria, of
chromalveolates by the enslaving of a
red alga (Cavalier-Smith, 1999, 2003;
Harper and Keeling, 2003) and in the
origin of the green plastids of
euglenoid (excavate) and chlorarachnean
(cercozoan) algae-a green algal cell
was enslaved either by the ancestral
cabozoan (arrow) or (less likely) twice
independently within excavates and
Cercozoa (asterisks) (Cavalier-Smith,
2003a). The upper thumbnail sketch
shows membrane topology in the
chimaeric cryptophytes (class
Cryptophyceae of the phylum Cryptista);
in the ancestral chromist the former
food vacuole membrane fused with the
rough endoplasmic reticulum placing the
enslaved cell within its lumen (red) to
yield the complex membrane topology
shown. The large host nucleus and the
tiny nucleomorph are shown in blue,
chloroplast green and mitochondrion
purple. In chlorarachneans (class
Chlorarachnea of phylum Cercozoa) the
former food vacuole membrane remained
topologically distinct from the ER to
become an epiplastid membrane and so
did not acquire ribosomes on its
surface, but their membrane topology is
otherwise similar to the cryptophytes.
The other sketches portray the four
major kinds of cell in the living world
and their membrane topology. The upper
ones show the contrasting ancestral
microtubular cytoskeleton (ciliary
roots, in red) of unikonts (a cone of
single microtubules attaching the
single centriole to the nucleus, blue)
and bikonts (two bands of microtubules
attached to the posterior centriole and
an anterior fan of microtubules
attached to the anterior centriole).
The lower ones show the single plasma
membrane of unibacteria (posibacteria
plus archaebacteria), which were
ancestral to eukaryotes and the double
envelope of negibacteria, which were
ancestral to mitochondria and
chloroplasts (which retained the outer
membrane, red).
source: http://aob.oxfordjournals.org/cg
i/content/full/95/1/147/FIG2
[2] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group.
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
now.
[1] FIG. 2. The tree of life based on
molecular, ultrastructural and
palaeontological evidence. Contrary to
widespread assumptions, the root is
among the eubacteria, probably within
the double-enveloped Negibacteria, not
between eubacteria and archaebacteria
(Cavalier-Smith, 2002b); it may lie
between Eobacteria and other
Negibacteria (Cavalier-Smith, 2002b).
The position of the eukaryotic root has
been nearly as controversial, but is
less hard to establish: it probably
lies between unikonts and bikonts (Lang
et al., 2002; Stechmann and
Cavalier-Smith, 2002, 2003). For
clarity the basal eukaryotic kingdom
Protozoa is not labelled; it comprises
four major groups (alveolates, cabozoa,
Amoebozoa and Choanozoa) plus the small
bikont phylum Apusozoa of unclear
precise position; whether Heliozoa are
protozoa as shown or chromists is
uncertain (Cavalier-Smith, 2003b).
Symbiogenetic cell enslavement occurred
four or five times: in the origin of
mitochondria and chloroplasts from
different negibacteria, of
chromalveolates by the enslaving of a
red alga (Cavalier-Smith, 1999, 2003;
Harper and Keeling, 2003) and in the
origin of the green plastids of
euglenoid (excavate) and chlorarachnean
(cercozoan) algae-a green algal cell
was enslaved either by the ancestral
cabozoan (arrow) or (less likely) twice
independently within excavates and
Cercozoa (asterisks) (Cavalier-Smith,
2003a). The upper thumbnail sketch
shows membrane topology in the
chimaeric cryptophytes (class
Cryptophyceae of the phylum Cryptista);
in the ancestral chromist the former
food vacuole membrane fused with the
rough endoplasmic reticulum placing the
enslaved cell within its lumen (red) to
yield the complex membrane topology
shown. The large host nucleus and the
tiny nucleomorph are shown in blue,
chloroplast green and mitochondrion
purple. In chlorarachneans (class
Chlorarachnea of phylum Cercozoa) the
former food vacuole membrane remained
topologically distinct from the ER to
become an epiplastid membrane and so
did not acquire ribosomes on its
surface, but their membrane topology is
otherwise similar to the cryptophytes.
The other sketches portray the four
major kinds of cell in the living world
and their membrane topology. The upper
ones show the contrasting ancestral
microtubular cytoskeleton (ciliary
roots, in red) of unikonts (a cone of
single microtubules attaching the
single centriole to the nucleus, blue)
and bikonts (two bands of microtubules
attached to the posterior centriole and
an anterior fan of microtubules
attached to the anterior centriole).
The lower ones show the single plasma
membrane of unibacteria (posibacteria
plus archaebacteria), which were
ancestral to eukaryotes and the double
envelope of negibacteria, which were
ancestral to mitochondria and
chloroplasts (which retained the outer
membrane, red).
source: http://aob.oxfordjournals.org/cg
i/content/full/95/1/147/FIG2
[2] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group.
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
evolve now.
[1] FIG. 2. The tree of life based on
molecular, ultrastructural and
palaeontological evidence. Contrary to
widespread assumptions, the root is
among the eubacteria, probably within
the double-enveloped Negibacteria, not
between eubacteria and archaebacteria
(Cavalier-Smith, 2002b); it may lie
between Eobacteria and other
Negibacteria (Cavalier-Smith, 2002b).
The position of the eukaryotic root has
been nearly as controversial, but is
less hard to establish: it probably
lies between unikonts and bikonts (Lang
et al., 2002; Stechmann and
Cavalier-Smith, 2002, 2003). For
clarity the basal eukaryotic kingdom
Protozoa is not labelled; it comprises
four major groups (alveolates, cabozoa,
Amoebozoa and Choanozoa) plus the small
bikont phylum Apusozoa of unclear
precise position; whether Heliozoa are
protozoa as shown or chromists is
uncertain (Cavalier-Smith, 2003b).
Symbiogenetic cell enslavement occurred
four or five times: in the origin of
mitochondria and chloroplasts from
different negibacteria, of
chromalveolates by the enslaving of a
red alga (Cavalier-Smith, 1999, 2003;
Harper and Keeling, 2003) and in the
origin of the green plastids of
euglenoid (excavate) and chlorarachnean
(cercozoan) algae-a green algal cell
was enslaved either by the ancestral
cabozoan (arrow) or (less likely) twice
independently within excavates and
Cercozoa (asterisks) (Cavalier-Smith,
2003a). The upper thumbnail sketch
shows membrane topology in the
chimaeric cryptophytes (class
Cryptophyceae of the phylum Cryptista);
in the ancestral chromist the former
food vacuole membrane fused with the
rough endoplasmic reticulum placing the
enslaved cell within its lumen (red) to
yield the complex membrane topology
shown. The large host nucleus and the
tiny nucleomorph are shown in blue,
chloroplast green and mitochondrion
purple. In chlorarachneans (class
Chlorarachnea of phylum Cercozoa) the
former food vacuole membrane remained
topologically distinct from the ER to
become an epiplastid membrane and so
did not acquire ribosomes on its
surface, but their membrane topology is
otherwise similar to the cryptophytes.
The other sketches portray the four
major kinds of cell in the living world
and their membrane topology. The upper
ones show the contrasting ancestral
microtubular cytoskeleton (ciliary
roots, in red) of unikonts (a cone of
single microtubules attaching the
single centriole to the nucleus, blue)
and bikonts (two bands of microtubules
attached to the posterior centriole and
an anterior fan of microtubules
attached to the anterior centriole).
The lower ones show the single plasma
membrane of unibacteria (posibacteria
plus archaebacteria), which were
ancestral to eukaryotes and the double
envelope of negibacteria, which were
ancestral to mitochondria and
chloroplasts (which retained the outer
membrane, red).
source: http://aob.oxfordjournals.org/cg
i/content/full/95/1/147/FIG2
[2] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group.
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
evolve now.
[1] FIG. 2. The tree of life based on
molecular, ultrastructural and
palaeontological evidence. Contrary to
widespread assumptions, the root is
among the eubacteria, probably within
the double-enveloped Negibacteria, not
between eubacteria and archaebacteria
(Cavalier-Smith, 2002b); it may lie
between Eobacteria and other
Negibacteria (Cavalier-Smith, 2002b).
The position of the eukaryotic root has
been nearly as controversial, but is
less hard to establish: it probably
lies between unikonts and bikonts (Lang
et al., 2002; Stechmann and
Cavalier-Smith, 2002, 2003). For
clarity the basal eukaryotic kingdom
Protozoa is not labelled; it comprises
four major groups (alveolates, cabozoa,
Amoebozoa and Choanozoa) plus the small
bikont phylum Apusozoa of unclear
precise position; whether Heliozoa are
protozoa as shown or chromists is
uncertain (Cavalier-Smith, 2003b).
Symbiogenetic cell enslavement occurred
four or five times: in the origin of
mitochondria and chloroplasts from
different negibacteria, of
chromalveolates by the enslaving of a
red alga (Cavalier-Smith, 1999, 2003;
Harper and Keeling, 2003) and in the
origin of the green plastids of
euglenoid (excavate) and chlorarachnean
(cercozoan) algae-a green algal cell
was enslaved either by the ancestral
cabozoan (arrow) or (less likely) twice
independently within excavates and
Cercozoa (asterisks) (Cavalier-Smith,
2003a). The upper thumbnail sketch
shows membrane topology in the
chimaeric cryptophytes (class
Cryptophyceae of the phylum Cryptista);
in the ancestral chromist the former
food vacuole membrane fused with the
rough endoplasmic reticulum placing the
enslaved cell within its lumen (red) to
yield the complex membrane topology
shown. The large host nucleus and the
tiny nucleomorph are shown in blue,
chloroplast green and mitochondrion
purple. In chlorarachneans (class
Chlorarachnea of phylum Cercozoa) the
former food vacuole membrane remained
topologically distinct from the ER to
become an epiplastid membrane and so
did not acquire ribosomes on its
surface, but their membrane topology is
otherwise similar to the cryptophytes.
The other sketches portray the four
major kinds of cell in the living world
and their membrane topology. The upper
ones show the contrasting ancestral
microtubular cytoskeleton (ciliary
roots, in red) of unikonts (a cone of
single microtubules attaching the
single centriole to the nucleus, blue)
and bikonts (two bands of microtubules
attached to the posterior centriole and
an anterior fan of microtubules
attached to the anterior centriole).
The lower ones show the single plasma
membrane of unibacteria (posibacteria
plus archaebacteria), which were
ancestral to eukaryotes and the double
envelope of negibacteria, which were
ancestral to mitochondria and
chloroplasts (which retained the outer
membrane, red).
source: http://aob.oxfordjournals.org/cg
i/content/full/95/1/147/FIG2
[2] Fig. 1. A consensus phylogeny of
eukaryotes. The vast majority of
characterized eukaryotes, with the
notable exception of major subgroups of
amoebae, can now be assigned to one of
eight major groups. Opisthokonts (basal
flagellum) have a single basal
flagellum on reproductive cells and
flat mitochondrial cristae (most
eukaryotes have tubular ones).
Eukaryotic photosynthesis originated in
Plants; theirs are the only plastids
with just two outer membranes.
Heterokonts (different flagellae) have
a unique flagellum decorated with
hollow tripartite hairs (stramenopiles)
and, usually, a second plain one.
Cercozoans are amoebae with filose
pseudopodia, often living with in tests
(hard outer shells), some very
elaborate (foraminiferans). Amoebozoa
are mostly naked amoebae (lacking
tests), often with lobose pseudopodia
for at least part of their life cycle.
Alveolates have systems of cortical
alveoli directly beneath their plasma
membranes. Discicristates have discoid
mitochondrial cristae and, in some
cases, a deep (excavated) ventral
feeding groove. Amitochondrial
excavates lack substantial molecular
phylogenetic support, but most have an
excavated ventral feeding groove, and
all lack mitochondria. The tree shown
is based on a consensus of molecular
(1-4) and ultrastructural (16, 17) data
and includes a rough indication of new
ciPCR ''taxa'' (broken black lines)
(7-11). An asterisk preceding the taxon
name indicates probable paraphyletic
group.
source: http://www.sciencemag.org/cgi/co
ntent/full/300/5626/1703
fossils.
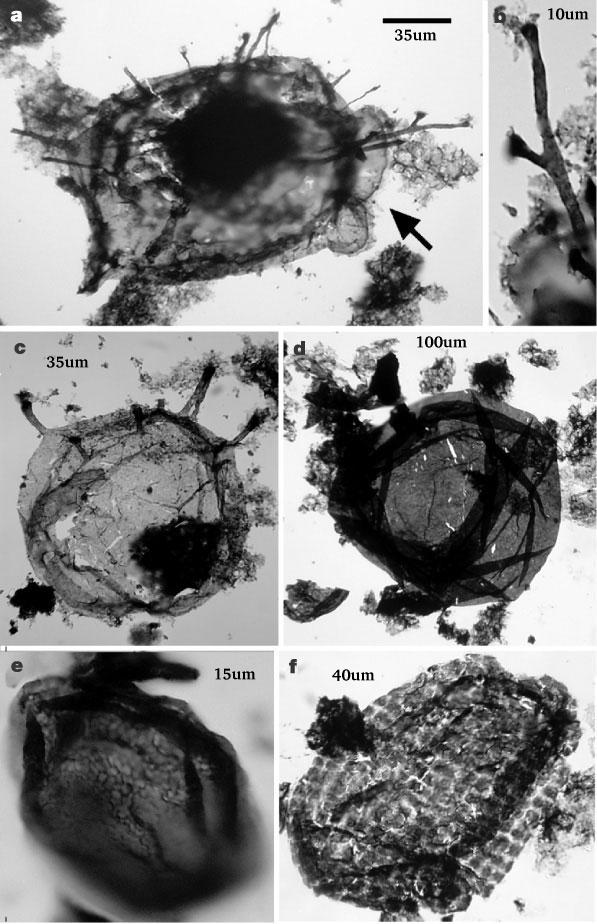
[1] Figure 1 Protistan microfossils
from the Roper Group. a, c, Tappania
plana, showing asymmetrically
distributed processes and bulbous
protrusions (arrow in a). b, detail of
a, showing dichotomously branching
process. d, Valeria lophostriata. e,
Dictyosphaera sp. f, Satka favosa. The
scale bar in a is 35 µm for a and c;
10 µm for b; 100 µm for d; 15 µm for
e; and 40 µm for f.
source: Nature 412
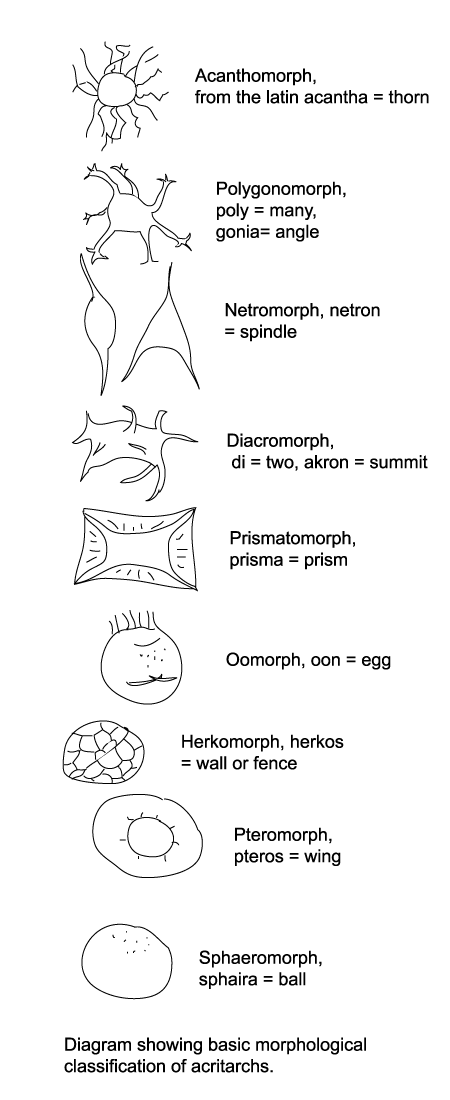
[2] Diagram showing basic
morphological classification of
acritarchs. COPYRIGHTED
source: http://www.ucl.ac.uk/GeolSci/mic
ropal/acritarch.html
fossil Grypania spiralis (an alga 10 cm
long) from BIF in Michigan. Oldest
algae fossil.
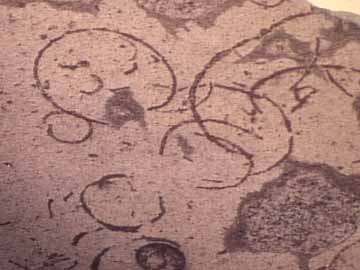
source: file:/root/web/Grypania_spiralis
_wmel0000.htm
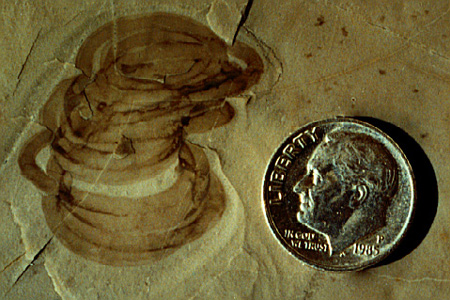
source: http://www.peripatus.gen.nz/pale
{kind=link}
ontology/lrgGrypaniaspiralis.jpg
Rocks.

source:
relationship with cyanobacteria, which
form plastids (chloroplasts). Like
mitochondria, these organelles copy
themselves and are not made by the cell
DNA.
to glaucophytes) evolves.
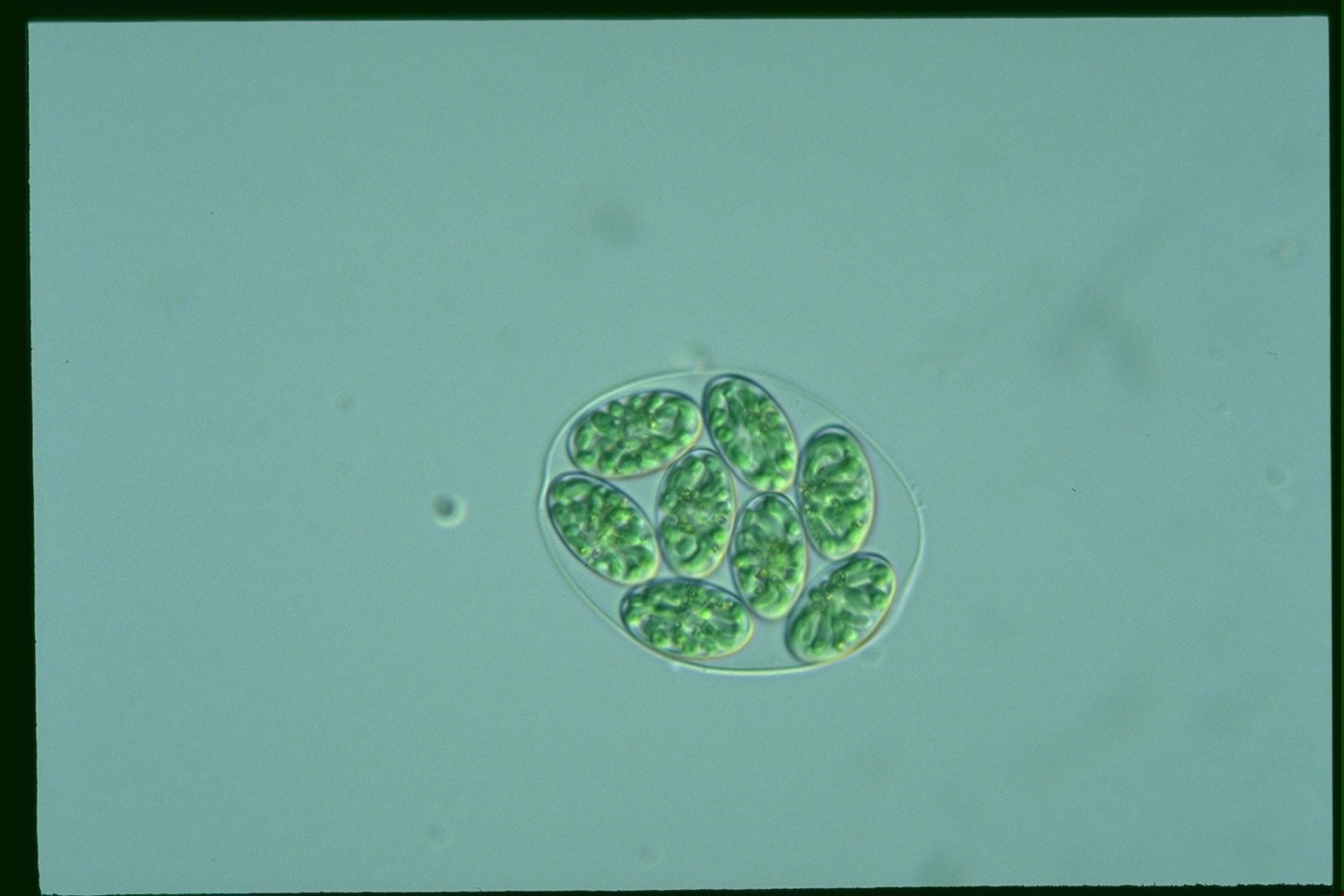
[1] ? COPYRIGHTED
source: http://protist.i.hosei.ac.jp/PDB
3/PCD3711/htmls/86.html

[2] ? COPYRIGHTED
source: http://protist.i.hosei.ac.jp/PDB
/Images/Others/Glaucocystis/
evolve from Unikonts (ancestrally only
one cilium). Opisthokonts have flat
mitochondrial cristae and go on to form
the Animal and Fungi kingdoms.

[1] cavalier-smith diagram COPYRIGHTED
source: cavalier_jmolevol_2003_56_540-56
3.pdf
[2] Figure 1. Phylogenetic hypothesis
of the eukaryotic lineage based on
ultrastructural and molecular data.
Organisms are divided into three main
groups distinguished by mitochondrial
cristal shape (either discoidal,
flattened or tubular). Unbroken lines
indicate phylogenetic relationships
that are firmly supported by available
data; broken lines indicate
uncertainties in phylogenetic
placement, resolution of which will
require additional data. Color coding
of organismal genus names indicates
mitochondrial genomes that have been
completely (Table 1), almost completely
(Jakoba, Naegleria and
Thraustochytrium) or partially (*)
sequenced by the OGMP (red), the FMGP
(black) or other groups (green). Names
in blue indicate those species whose
mtDNAs are currently being sequenced by
the OGMP or are future candidates for
complete sequencing. Amitochondriate
retortamonads are positioned at the
base of the tree, with broken arrows
denoting the endosymbiotic origin(s) of
mitochondria from a Rickettsia-like
eubacterium. Macrophar.,
Macropharyngomonas.
source: unknown
evolve now.
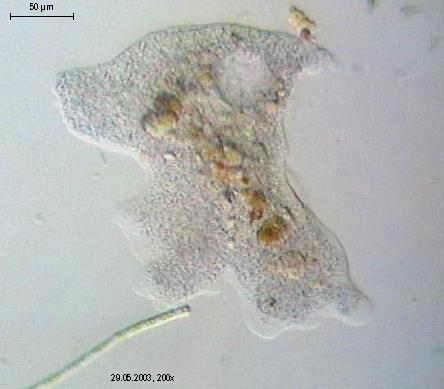
[1] SUBPHYLUM Lobosa CLASS Amoebaea
Chaos diffluens, an amoeba. Photo
released by Dr. Ralf Wagner.
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Chaos_diffluens.jpg
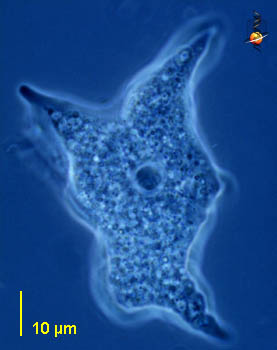
[2] CLASS Amoebaea Mayorella
(may-or -ell-a) a medium sized
free-living naked amoeba with conical
pseudopodia. Central body is the
nucleus. Phase contrast. This picture
was taken by David Patterson of
material from Limulus-ridden sediments
at Plum Island (Massachusetts USA) in
spring and summer, 2001. NONCOMMERCIAL
USE
source: http://microscope.mbl.edu/script
s/microscope.php?func=imgDetail&imageID=
515
Phlya Chlorophyta (volvox, sea lettuce)
and Charophyta (Spirogyra) evolve.
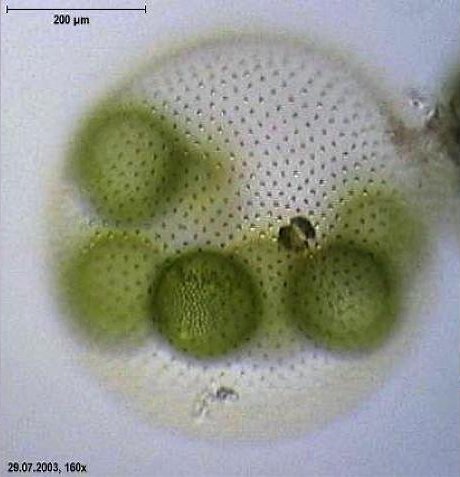
[1] Micrograph of Volvox aureus.
Copyright held by Dr. Ralf Wagner,
uploaded to German Wikipedia under
GFDL. Permission is granted to copy,
distribute and/or modify this document
under the terms of the GNU Free
Documentation License, Version 1.2 or
any later version published by the Free
Software Foundation; with no Invariant
Sections, no Front-Cover Texts, and no
Back-Cover Texts. Subject to
disclaimers.
source: http://en.wikipedia.org/wiki/Vol
vox

[2] Photo of green algal growth
(Enteromorpha sp.) on rocky areas of
the ocean intertidal shore, indicating
a nearby nutrient source (in this case
land runoff). Photographed by Eric
Guinther near Kahuku, O'ahu,
Hawai'i. GFDL Permission is granted
to copy, distribute and/or modify this
document under the terms of the GNU
Free Documentation License, Version 1.2
or any later version published by the
Free Software Foundation; with no
Invariant Sections, no Front-Cover
Texts, and no Back-Cover Texts Subject
to disclaimers
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Intertidal_greenalgae.jpg

[1] Close-up of a red alga (Genus?
Laurencia), Class Florideophyceae,
Order=? a marine seaweed from Hawaii.
GNU
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Laurencia.jpg

[2] Bangia atropurpurea Profile:
unbranched filaments in tufts. Often
forming dense fringes in the spalsh
zone. Uniseriate at base, multiseriate
above with protoplasts separate in a
firm gelatinous sheath. Stellate
chloroplasts. US NOAA PD
source: http://www.glerl.noaa.gov/seagra
nt/GLWL/Algae/Rhodophyta/Cards/Bangia.ht
ml
is enslaved by a chromealveolate
eukaryote to form a plastid in the
chromealveolate. This kind of plastid
is presumably inherited by all other
chromalveolates (brown algae, diatoms,
water molds, Dinoflagellata,
Apicomplexa, ciliates) that have
plastids.
(red algae) fossils (Bangiomorpha
pubescens) from Hunting Formation,
Somerset Island, arctic Canada.

[1] get images from Life on a Young
Planet, Knoll
source: Science 1990 vol 250
Butterfield N. J. A. H. Knoll K. Swett
1990 A bangiophyte red alga from the
Proterozoic of Arctic Canada. Science
250: 104-107[ISI][Medline]

[2] Figure 2. Griffithsia pacifica
(Florideophyceae). Electron micrograph
showing cytoplasm with numerous
chloroplasts (C) and starch (S). Starch
is the photosynthetic reserve and is
deposited free in the cytoplasm.
source: (American Journal of Botany.
2004;91:1494-1507.)
"Microsporidia" evolves.
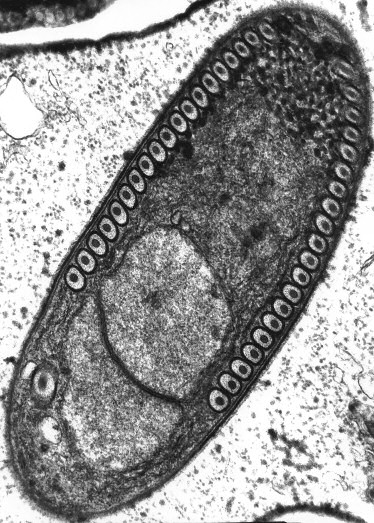
[1] Sporoblast of the Microsporidium
Fibrillanosema crangonycis. Electron
micrograph taken by Leon White. GNU
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Fibrillanosema_spore.jpg
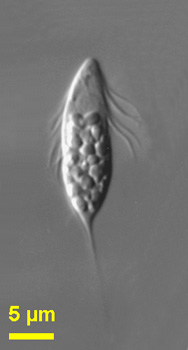
[2] Spironema
multiciliatum Spironema:
Octosporoblastic sporogony producing
horseshoe-shaped monokaryotic spores in
sporophorous vesicles; monomorphic,
diplokaryotic and monokaryotic;
merogony - last generation merozoites
are diplokaryotic; sporogony - initial
division of the sporont nuclei is
meiotic as indicated by the occurrence
of synaptonemal complexes; spores are
horse-shoe-shaped, with swollen ends in
T. variabilis and have one elongate
nucleus; exospore with three layers,
endospore is of medium thickness;
polaroplast composed of two lamellar
parts, an anterior part of closely
packed lamellae and a posterior part of
wider compartments; polar tube is
isofilar and forms, in the posterior
quarter of the spore, 3-4 coils in a
single rank (T. variabilis) or 8-10
coils in a single rank (T. chironomi);
type species Toxoglugea vibrio in
adipose tissue of larvae of Ceratopogon
sp. (Diptera, Ceratopogonidae).
Spironema (spire-oh-knee-ma)
multiciliatum Klebs, 1893. Cells are
lanceolate, relatively flattened and
flexible. The cells have a spiral
groove, long kinetics and a tail, which
tapers posteriorly, and are about 15 -
21 microns without the tail. The
nucleus is located anteriorly or near
the centre of the cell. When the cells
are squashed, the cells are more
flexible. Food materials are seen under
the cell surface. Rarely observed.
This picture was taken by Won Je Lee
using conventional photographic film
using a Zeiss Axiophot microscope of
material collected in marine sediments
of Botany Bay (Sydney, Australia). The
image description refers to material
from Botany Bay. NONCOMMERCIAL USE
source: http://microscope.mbl.edu/script
s/microscope.php?func=imgDetail&imageID=
3928
evolves.

[1] Chytrids (Chytridiomycota): The
Primitive Fungi These fungi are
mostly aquatic, are notable for having
a flagella on the cells (a flagella is
a tail, somewhat like a tail on a sperm
or a pollywog), and are thought to be
the most primitive type of
fungi. actual photo comes
from: http://www.csupomona.edu/~jcclark
/classes/bot125/resource/graphics/chy_al
l_sph.html
source: http://www.davidlnelson.md/Cazad
ero/Fungi.htm
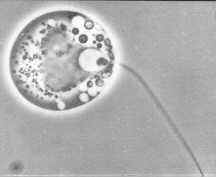
[2] Chytridiomycota - Blastocladiales
- zoospore of Allomyces (phase contrast
illumination) X 2000
source: http://www.mycolog.com/chapter2b
.htm
(Mesomycetozoea/DRIPs,
Choanoflagellates) evolves.
(DRIPs) evolve.
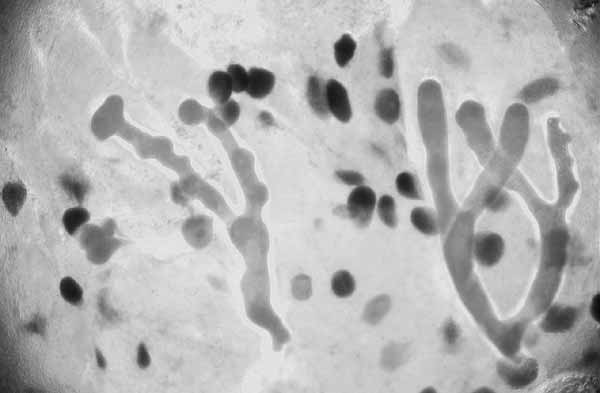
[1] Ichthyophonus, a fungus-like
protistan that occurs in high
prevalence in Pacific Ocean perch
(Sebastes aultus) and yellowtail
rockfish (Sebastes flavedus). Note the
parasite forms branching hyphae-like
structures. Ichthyophonus hoferi has
caused massive mortalities in herring
in the Atlantic ocean, and has recently
been reported to cause disease in wild
Pacific herring from Washington through
Alaska. COPYRIGHTED EDU
source: http://oregonstate.edu/dept/salm
{kind=link}
on/projects/images/16Ichthyophonus.jpg
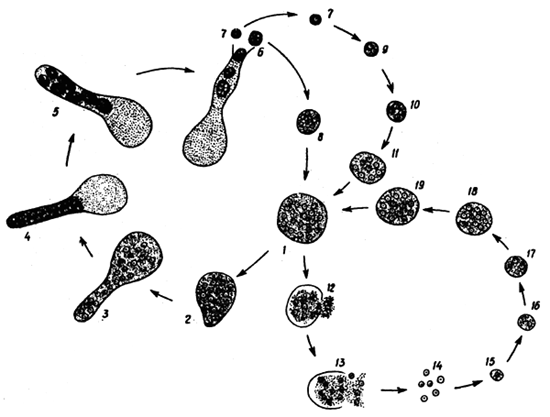
[2] Microscopic appearence of the
organism is dependent on its stage of
development. The stages include (1)
spore at ''resting'' stage, (2)
germinating spore, (3) hyphal
stage. It is believed that there are
two forms of Ichthyophonus, both
belonging to one genus. One of them is
known as the ''salmon'' form, occuring
in freshwater and cold-preferring sea
fishes: this form is characterized by
its ability to produce long tubulose
germ hyphae. The other is called the
''aquarium fish'' form, typical of the
tropical freshwater fishes. This form
is completely devoid of hyphae.
Developmental cycle of Ichthyophonus
hoferi: 1-5 - development of
''daughter'' spores, 7-11 - development
of resting spore from the ''daughter''
spore, 12-19 - development of resting
spore by fragmentation. COPYRIGHTED
source: http://www.fao.org/docrep/field/
003/AC160E/AC160E02.htm
evolve in unicellular eukaryote living
objects. This is the first proto eye.
"Choanoflagellates" and "Acanthoecida"
evolve.

[1] DOMAIN Eukaryota - eukaryotes
KINGDOM Protozoa (Goldfuss, 1818) R.
Owen, 1858 - protozoa SUBKINGDOM
Sarcomastigota (means=?) PHYLUM
Choanozoa CLASS
Choanoflagellatea (Choanoflagellates
and Acanthoecida) ORDER
Acanthoecida Saepicula: Cells
solitary, lorica funnel-shaped, 2
chambers delimited by a waist;
constructed of rod-shaped costal
strips; posterior chamber obconical
with 2 series of costae located more or
less regularly around chamber, one
series almost parallel to the long axis
of cell and second series almost
perpendicular to long axis; anterior
chamber formed by ring of equally
spaced longitudinal costae surmounted
by single transverse costa; marine
This image is based on a drawing
provided by Won Je Lee. NONCOMMERCIAL
USE
source: http://microscope.mbl.edu/script
s/microscope.php?func=imgDetail&imageID=
3229

[2] Choanoeca: Cells solitary with
distinct, firm flask-shaped theca more
or less closely investing protoplast,
with short pedicel; collar relatively
long, widely expanded; flagellum absent
in adult, but produced prior to cell
division for locomotory use by juvenile
cell; in marine and brackish habitats,
frequently attached to filamentous
algae and hydrozoa Choanoeca
(ko-an-o-eek-a), an unusual loricate
collar flagellate (choanoflagellate) in
that the usual form is without a
flagellum. Flagellated motile stage is
occasionally produced. Widely dispersed
pseudopodial elements of the collar are
evident in this image. Differential
interference contrast. This picture
was taken by David Patterson and Aimlee
Laderman of material collected from a
freshwater Atantic white cedar swamp at
Cumloden near Woods Hole in
Massachusetts, USA in spring and
summer, 2001. NONCOMMERCIAL USE
source: http://microscope.mbl.edu/script
s/microscope.php?func=imgDetail&imageID=
170
multicellularity evolves, where a
zygote produces differentiated cells
that stick together to form one
organism.
evolve. Metazoans are multicellular,
but their cells perform different
functions and originate from one
cell(?). This is`also the beginning of
the Animal Subkingdom "Radiata",
species with radial symmetry. These are
the sponges. There are only 3 kinds of
metazoans: sponges, cnidarians, and
bilaterians (which include all insects
and vertibrates). Sponges are the
first organisms whose DNA codes for
more than one kind of cell. Sponges
have 3 different cell types. Some
cells form a body wall, some digest
food, some form a skeletal frame.

[1]
source: http://www.museums.org.za/bio/me
tazoa.htm

[2]
source: http://www.museums.org.za/bio/me
tazoa.htm
evolve. These genes regulate the
building of major body parts.
division "Zygomycota" (bread molds, pin
molds, microsporidia,...) evolving now.
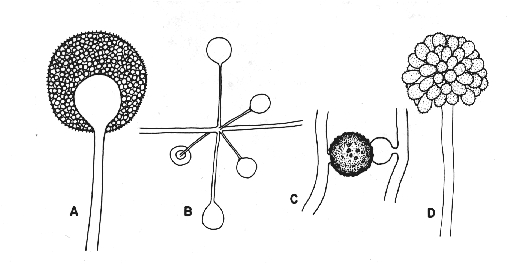
[1] Figure 2. Zygomycota A: sporangia
of Mucor sp. B: whorl of sporangia of
Absidia sp. C: zygospore of
Zygorhynchus sp. D: sporangiophore and
sporangiola of Cunninghamella sp.
source: http://www.botany.utoronto.ca/Re
searchLabs/MallochLab/Malloch/Moulds/Cla
ssification.html
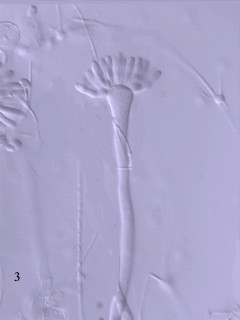
[2] Figure 3. Syncephalis, a member
of the Zygomycota parasitic on other
Zygomycota
source: http://www.botany.utoronto.ca/Re
searchLabs/MallochLab/Malloch/Moulds/Cla
ssification.html
jellies) evolves.
division "Glomeromycota" (Arbuscular
mycorrhizal fungi) evolving now.

[1] germinating Gigaspora decipiens
source: http://pages.unibas.ch/bothebel/
people/redecker/ff/glomero.htm
[2] Archaeospora leptoticha spores
source: http://pages.unibas.ch/bothebel/
people/redecker/ff/glomero.htm
jellyfish evolves. Jellyfish have
photon detecting cells and a lens made
of ?.
the phylum "Basidiomycota" (most
mushrooms, rusts, club fungi) evolve.

[1] Amanita muscaria
(Homobasidiomycetes)
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Agaricales.jpg

[2] Basidiomycete Life Cycle tjv
source: http://botit.botany.wisc.edu/ima
ges/332/Basidiomycota/General_basidio/Ba
sidiomycete_Life_Cycle_tjv.php?highres=t
rue
"Ascomycota" (yeasts, truffles,
Penicillium, morels, sac fungi)
evolves.

[1] white truffle
cutted photographed by
myself GNU head Permission is
granted to copy, distribute and/or
modify this document under the terms of
the GNU Free Documentation License,
Version 1.2 or any later version
published by the Free Software
Foundation; with no Invariant Sections,
no Front-Cover Texts, and no Back-Cover
Texts. A copy of the license is
included in the section entitled ''Text
of the GNU Free Documentation
License.''
source: http://upload.wikimedia.org/wiki
{kind=link}
pedia/commons/f/fd/Truffle_washed_and_cu
tted.jpg
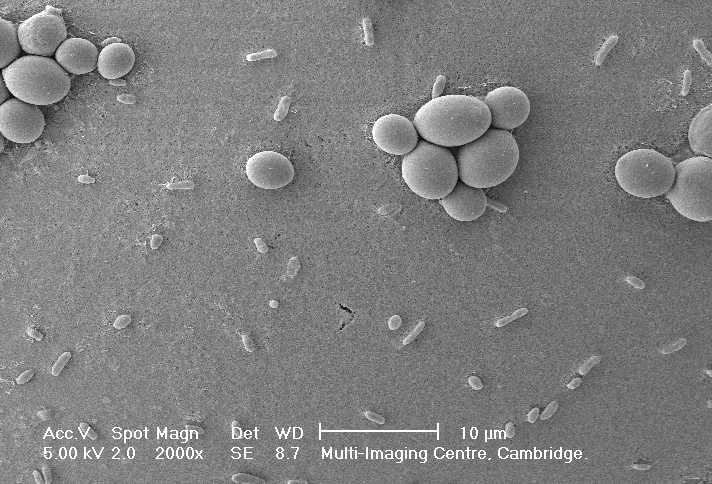
[2] EColi-Scerevisiae.jpg (50KB, MIME
type: image/jpeg) Wikimedia Commons
logo This is a file from the Wikimedia
Commons. The description on its
description page there is shown
below. Escherichia coli (little
forms) & Saccharomyces cerevisiae (big
forms) by MEB Public domain This file
has been released into the public
domain by the copyright holder, its
copyright has expired, or it is
ineligible for copyright. This applies
worldwide. brewer's yeast/baker's
yeast
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:EColi-Scerevisiae.jpg
largest and second largest lines of
Fungi (Ascomycota and Basidiomycota)
splitting now.
symmetry) species evolves. Animal
phylum Acoelomorpha (acoela flat worms
and nemertodermatida) evolves.
This
begins the Subkingdom "Bilateria".
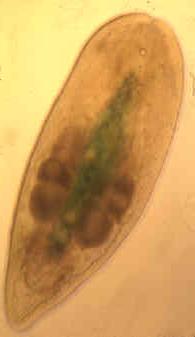
[1] Convoluta pulchra Smith and Bush
1991, a typical mud-inhabiting acoel
that feeds on diatoms
source: ?
evolve at this time. Protostomes
include the 3 infrakingdoms Ecdysozoa
(a variety of worms and the arthropods
{a huge group including all insects and
crustaceans}), Platyzoa (rotifers and
flatworms), and Lophotrochozoa
(brachiopods {clams}, molluscs
{snails}, and a variety of worms).
Doushantuo formation in China.
Vernanimalcula, 178 um in length, from
Doushantuo Formation, China. First
fossil of organism with bilateral
symmetry, mouth, digestive track, gut
and anus.
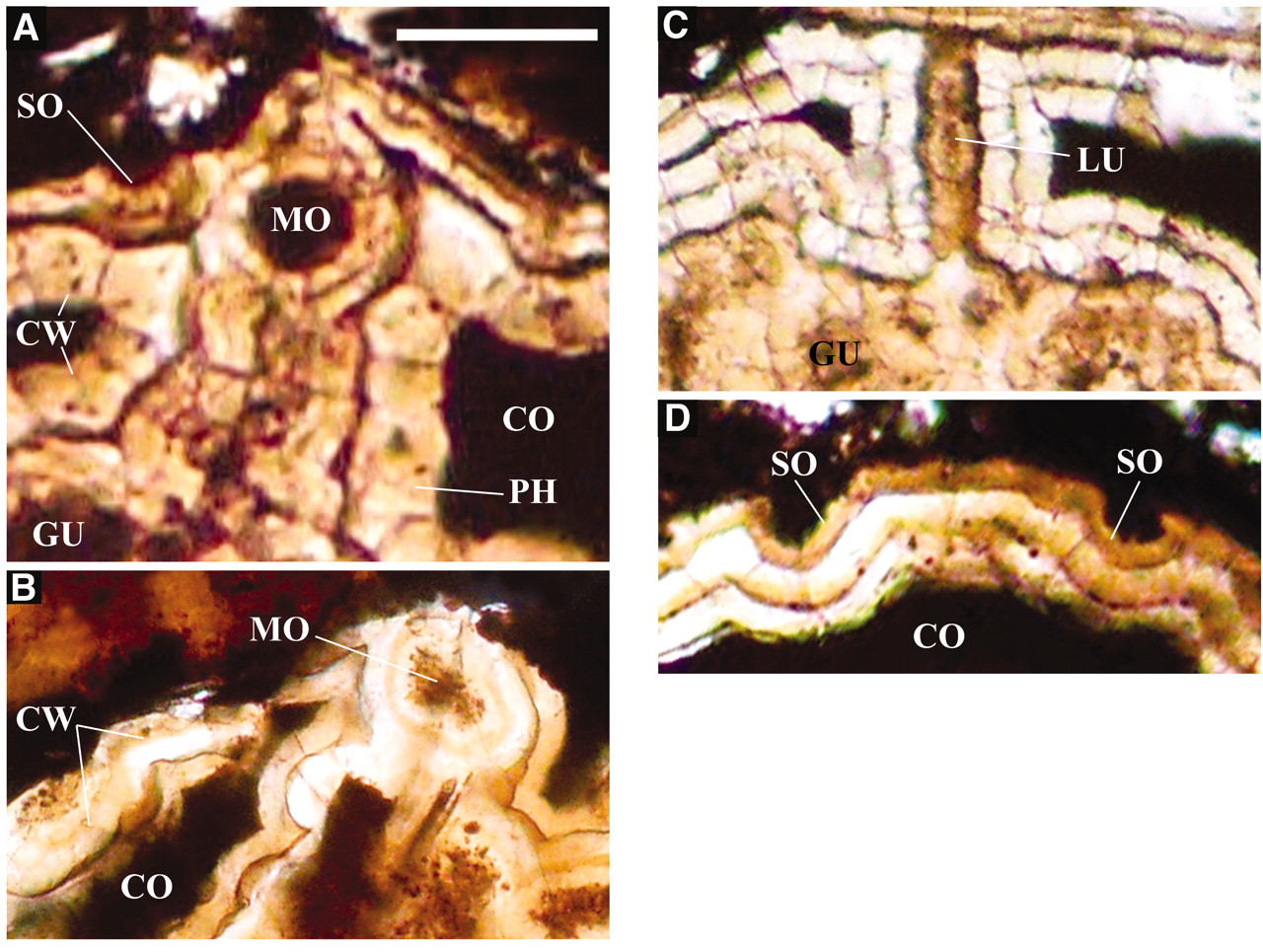
[1] Fig. 2. Close-up images of
prominent anatomical features of
Vernanimalcula guizhouena. The scale
bar represents 18 µm in (A), 32 µm in
(B), 24 µm in (C), and 28 µm in (D).
SO, sensory organ, i.e., external pit;
LU, lumen; PH, pharynx; MO, mouth; CO,
coelomic lumen; CW, mesodermal coelomic
wall; GU, gut. (A) Detail of collared
mouth, multilayered pharynx, and one
anterior surface pit. In this image,
which is from the holotype specimen
(Fig. 1A), the floor of the pit can be
seen to be composed of a specialized
concave layer. Note the coelomic wall,
which here as elsewhere in these
specimens has a thickness of about 5 to
6 µm. (B) Mouth of a fourth specimen,
Q3105, displaying collared mouth and
pharynx, ventral view. (C) Lumen of
pharynx from a fifth specimen, X10419,
secondarily encrusted but revealing
morphology of opening of pharynx into
gut similar to that seen in the
specimens shown in Fig. 1. (D) Close-up
of spaced external pits, interpreted as
possible sensory organs, from the same
specimen as shown in Fig. 1B [compare
(A)].
source: http://www.sciencemag.org/cgi/co
ntent/full/sci;305/5681/218
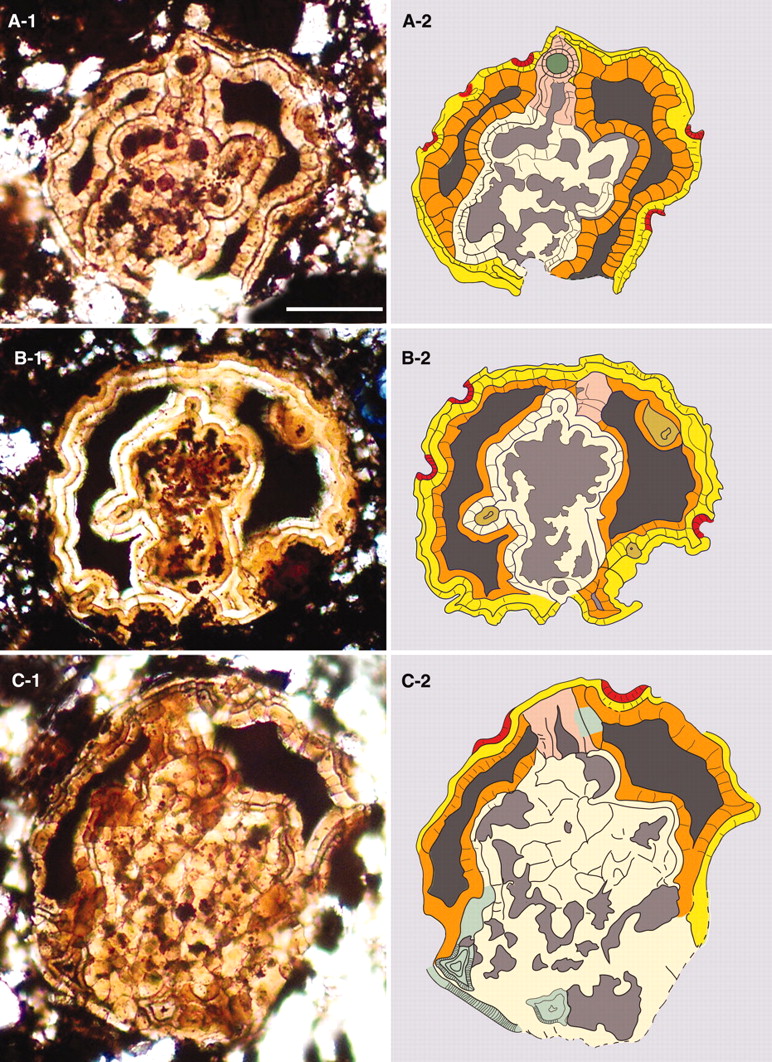
[2] Fig. 1. Images of three
different, fairly well preserved
specimens of the bilaterally organized
fossil animal Vernanimalcula
guizhouena. Left panels show digitally
recorded, transmitted light images of
sections about 50 µm thick, which had
been ground from larger rock samples,
mounted on slides, and viewed through a
light microscope. Right panels show
color-coded representations of the
images on the left. These were prepared
by digital image overlay. Yellow,
external ectodermal layer; ochre,
coelomic mesodermal layer; red, surface
pits; mauve, pharynx; light tan,
endodermal wall of gut; gray-green,
lumen of mouth; dark gray, paired
coelomic cavities; lighter gray, lumen
of gut; brown, ''gland-like''
structures, with central lumen (B);
light green, mineral inclusions (C).
The scale bar represents 40 µm in (A),
55 µm in (B), and 46 µm in (C). (A)
Holotype specimen, X00305, slightly
tilted, almost complete ventral level
coronal section, passing through the
ventrally located mouth. (B) Coronal
section of second specimen, X08981,
passing through dorsal wall of pharynx
and displaying complete A-P length of
digestive tract, including posterior
end [not visible in (A)]. (C) Tilted
coronal section of third specimen,
X10475, possibly slightly squashed,
passing through dorsal wall of pharynx
and through the dorsal wall of the gut.
For dimensions, see Table 1.
source:
evolves. Ecdysozoa are animals that
molt (lose their outer skins) as they
grow.
Ecdysozoa include:
the Phylum "Chaetognatha"
(Arrow Worms),
the Superphylum
"Aschelminthes", containing the 5
Phlya:
"Kinorhyncha" (kinorhynchs)
"Loricifera"
(loriciferans)
"Nematoda" (round worms)
"Nematomorpha"
(horsehair worms),
"Priapulida" (priapulids)
the
Superphlyum "Panarthropoda" containing
the 3 Phyla:
"Arthropoda" (arthropods:
insects, shell fish)
"Onychophora"
(onychophorans)
"Tardigrada" (tardigrades)
nervous system evolves in bilaterians.
in early bilaterians.
beginning of the Subkingdom
Deuterostomia and Infrakingdom
"Coelomopora" (Ambulacraria) with the
two Phyla "Hemichordata" (acorn worms)
and "Echinodermata" (sea cucumbers, sea
urchins, starfish).
(Arrow Worms) evolves.
Hemichordonia (acorn worms) evolves.
Echinodermata (sea cucumbers, sea
urchins, sand dollars, star fish)
evolves.
blood cells evolve in bilaterian worms.
Superphylum Gnathifera {gnathiferans},
Phylum Gastrotricha {gastrotrichs}, and
Phylum Platyhelminthes {flatworms})
evolve.
evolves. Chordata is a very large
group that contains all fish,
amphibians, reptiles and mammals.
Tunicata (tunicates {sea squirts})
evolves.
oxygen from water through a digestive
system, evolves in segmented worms.
Ashelminthes (round worms, horsehair
worms, priapulids) and Pananthropoda
(arthropods, onychophorans,
tardigrades) separate.
Cephalochordata (lancelets) evolves.
This is the first fish.
"Ashelminthes" evolves. This includes
the 5 Phyla:
Kinorhyncha (kinorhynchs),
Loricifera (loriciferans),
Nematoda (round worms),
Nematomorpha
(horsehair worms),
Priapulida (priapulids).
evolves. This includes the 5 Phyla:
Gna
thostomulida (gnathostomulids),
Cycliophora
(cycliophorans),
Micrognathozoa,
Rotifera (rotifers),
Acanthocephala
(acanthocephalans).
Lophotrochozoa evolves. This includes
brachiopods, bryozoans, clams, squids
and octopuses (cephalopods), and
snails.
Lophophorata evolves. This includes
the two Phyla Phoronida (phoronids) and
Brachiopoda (brachiopods {clams,
oysters, muscles}).
(phoronids) evolves.
Brachiopoda (brachiopods {clams,
oysters, muscles}) evolves.
Entoprocta (entoprocts) evolves.
Phanerozoic Eon. End Proterozoic Era,
start Paleozoic Era.
(flatworms) and Gastrotricha
(gastrotrichs) evolve.
mybn).
(Bryozoans or moss animals) evolves.
Panarthropoda (Arthropods, Onychophora,
Tardigrada) evolves.
(insects, crustaceans) evolve.
(onychophorans) evolves.
(tardigrades) evolves.

[1] Aldanella may be mollusc, if
mollusc may be first known snail.
shell is 1.5 mm in diameter.
source: http://www.geology.ucdavis.edu/~
cowen/HistoryofLife/CH05images.html
Eutrochozoa (molluscs, ribbon, peanut,
spoon, and segmented worms) evolves.
evolve.

[1]
http://www.ucmp.berkeley.edu/porifera/ar
chaeo.html
source: http://www.ucmp.berkeley.edu/por
ifera/archaeo.html

[2]
http://www.geology.ucdavis.edu/~cowen/Hi
storyofLife/CH05images.html
source: http://www.geology.ucdavis.edu/~
cowen/HistoryofLife/CH05images.html
(ribbon worms) evolves.
(brachiopods, bryozoans, clams,
mussels, squids and octopuses
{cephalopods}, and snails) evolves.
(segmented worms) evolve.
Sipuncula (peanut worms) evolve.
Vertebrata evolves. This Subphylum
contains most fish, all amphibians,
reptiles, and mammals.
(agnatha) evolve.

[1] Figure 4 The Lower Cambrian
agnathan vertebrate Haikouichthys
ercaicunensis Luo, Hu & Shu gen. et sp.
nov. from Haikou, Yunnan. Specimen
HZ-f-12-127. a, Entire specimen,
anterior to the left; more posterior
region appears to fade out into
sediment, possibly representing decay
of body; attempts to excavate this area
were not successful. Scale bar
equivalent to 5 mm. b, Detail of
anterior to show putative gill bars,
possible elements of cranial
endoskeleton, and pericardic area;
scale bar equivalent to 5 mm. c,
Camera-lucida drawing of specimen to
show interpretation. Numbers 1-6
indicate units of the branchial basket
that are identified with some
confidence; ?A-?C refer to less secure
identifications. Two possible areas
representing the pericardic cavity are
indicated. To the anterior of ?C a
triangular area with patches of
diagenetic mineralization is one
possibility; a fainter region to the
posterior is the alternative location.
COPYRIGHTED
source: http://www.nature.com/nature/jou
rnal/v402/n6757/fig_tab/402042a0_F4.html
measured in Kopli quarry, Tallinn,
Estonia.
Burgess Shale.

[1] diagram
source: http://www.nmnh.si.edu/paleo/sha
le/pmarella.htm

[2] fossil
source: http://www.nmnh.si.edu/paleo/sha
le/pmarella.htm
end Cambrian period (543-490 mybn).
ancestor of all plants (Kingdom
Plantae) evolving at this time (in the
view that algae are protists and not
plants).
source: http://protist.i.hosei.ac.jp/PDB
3/PCD3711/htmls/86.html
source: http://protist.i.hosei.ac.jp/PDB
/Images/Others/Glaucocystis/
non-vascular plant and vascular plant
lines splitting now.
non-vascular plants (Bryophytes)
(Liverworts, Hornworts, Mosses)
evolving now.

[1] Phaeoceros laevis (L.) Prosk. gnu
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Anthoceros_levis.jpg

[2] Image of Phaeoceros (hornwort)
spores taken by J. Ziffer. public
domain
source: wiki
lampreys and hagfish lines separate.
land plants.
Gnathostomata) evolve. This large group
includes all jawed fish, all
amphibians, reptiles, and mammals.
Gnathostomata) Class Chondrichthyes
(cartilaginous fishes) evolve.
phylum includes all tunicates, fishes,
amphibians, reptiles, birds, and
mammals. The living chordate with the
oldest DNA design are tunicates.
end Ordovician period (490-443 mybn).
Ray-finned fishes (Subclass
Actinopterygii) evolve.
plants, Cooksonia.

[1] Cooksonia pertoni with three
sporangia. Height of the plant 2.5
cm Pridolian (Upper
Silurian) Shropshire, England.
COPYRIGHTED
source: http://www.xs4all.nl/~steurh/eng
cook/ecookwal.html

[2] Cooksonia pertoni, fossilised
plant COPYRIGHTED UK
source: http://owen.nhm.ac.uk/piclib/web
{kind=link}
images/0/0/900/936_sml.jpg
millipede Pneumodesmus.
mybn), end Silurian period (443-417
mybn).
oldest line of living vascular plants
from the Division "Lycophyta" evolving
now.

[1] Lycopodiella cernua (L.) Pic. Serm.
plant from windward O'ahu (Hawai'i)
taken in December 2003 by Eric Guinther
and released under the GNU Free
Documentation License. gnu
source: http://en.wikipedia.org/wiki/Lyc
ophyte

[2] Fossil trunk of Lepidodendron
aculeatum showing leaf scars gnu
source: http://en.wikipedia.org/wiki/Lep
idodendron
[1] Rhyniognatha hirsti. COPYRIGHTED
source: http://www.nhm.ac.uk/nature-onli
ne/earth/fossils/article-oldest-insect-f
ossil/the-oldest-fossil-insect-in-the-wo
rld.html
Chondrichthyes) Subclass
Subterbranchialia and Subclass
Elasmobranchii (shark-like fishes)
separate.
Superorder Holocephali (chimaeras: eg.
elephant fish) evolves.
line and the line that leads to Seed
Plants (Gymnosperms and Angiosperms)
separating now.
producing and Seed producing plant
lines separating now.
forests.
vertebrate.
(Plant Division "Pteridophyta")
evolving now.

[1] Ferns, Melbourne Botanical
Gardens gnu
source: http://en.wikipedia.org/wiki/Fer
n

[2] An Australian tree fern growing
on O'ahu, Hawai'i. Photographed by Eric
Guinther. A tree fern unrolling a new
frond GNU
source: same
ice age.
(354-290 mybn), end Devonian period
(417-354 mybn).
Superdivision Chondrostei (sturgeons
and paddlefish) evolves.
Infradivsion Cladistia (Bichirs)
evolves.
toads, Salamanders) evolve.
oldest living Gymnosperms from the
Plant Kingdom evolving now.

[1] Leaves and female cone of Cycas
revoluta GNU
source: http://en.wikipedia.org/wiki/Cyc
ad

[2] Leaves and male cone of Cycas
revoluta Photo of Cycas (sago cycad)
inflorescence, taken July 2001 by
User:Stan Shebs Cycas revoluta - male
plant GNU
source: same
Gymnosperms and Angiosperms lines
separating now.
(mayfly?).
result of the collision of Europe and
eastern North America.
This group, the Amniota, will
branch into the 3 major Classes:
Reptiles (Sauropsida), Birds (Aves),
and Mammals (Synapsida).
evolve.
evolve.
mybn), end Carboniferous period
(354-290 mybn).
diapsids (crocodiles) separate.
Infradivision Actinopteri evolves.
Infradivision Actinopteri, Gars evolve.
happens. This is the most devastating
mass extinction event in the history of
earth.
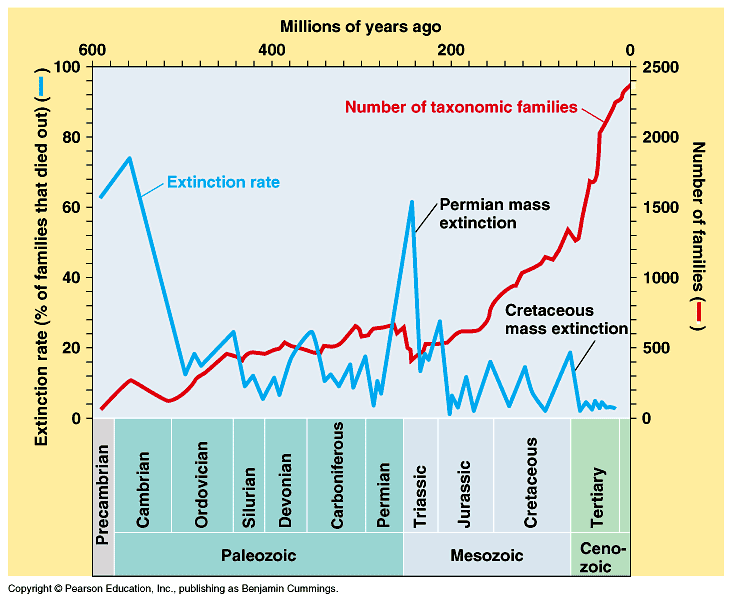
[1] Timeloine of mass extinctions.
COPYRIGHTED Benjamin Cummings.
source: http://io.uwinnipeg.ca/~simmons/
16cm05/1116/16macro.htm
Era.
mybn), end Permian period (290-248
mybn).
evolve.
Neopterygii evolves.
Subdivision Halecomorphi, Bow fish
(Amiiformes) evolve.
was found in South America.
herrings, anchovies, carp, minnows,
piranha, salmon, trout, pike, perch,
seahorse, cod) evolves.
lizards evolve.
evolve.
mybn), end Triassic period (248-206
mybn).
evolve.
because of climate (temperature?,
weather?) changes. Large outpourings
of lava from break-up of Pangea may
have caused climate change.
(shark-like fishes) divides into 2
divisions Squalea (rays, skates) and
Galeomorphii (great white, hammerhead,
nurse, sand tiger sharks).
evolve.
white, hammerhead, nurse, sand tiger
sharks) evolve.
Chromalveolates) fossils.
source: http://www.nature.com/news/2003/
{kind=link}
030217/images/diatom_180.jpg
source: http://www.ucmp.berkeley.edu/chr
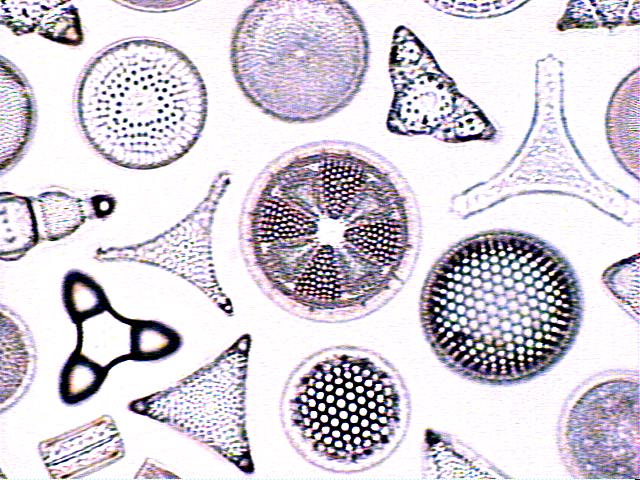{kind=link}
omista/diatoms/diatomdiverse.jpg
Monotremes lay eggs and are the
oldest warm blooded species of record.
ancient flowering plant (Angiosperm)
still alive, "Amborella" evolving now.

[1] Photo of Amborella trichopoda
(Amborellaceae; photo © Sangtae Kim).
source: http://tolweb.org/tree?group=ang
iosperms

[2] none
source: http://www.ucsc.edu/currents/99-
00/08-30/amborella.photo2.htm

[1] Archaeopteryx siemensii HMN
1880/81 (Berlin) COPYRIGHTED EDU
source: http://www.oucom.ohiou.edu/dbms-
witmer/dinoskulls02.htm

[2] Archaeopteryx sp. JM 2257
(Eichstätt) COPYRIGHTED EDU
source: http://www.oucom.ohiou.edu/dbms-
witmer/dinoskulls02.htm
Unlike Archaeopteryx, Confuciusornis
had no teeth.

[1] Confuciusornis
source: http://www.ucmp.berkeley.edu/dia
{kind=link}
psids/birds/confuciusornislg.jpg
branch of mammals) evolve.

[1] Archaefructus liaoningensis. The
leaf-like structures on the stem of
this 140 million year old fossil are
pods containing the seeds, a
characteristic unique to flowering
plants. Credit: University of Florida.
PD?
source: http://science.nasa.gov/headline
s/y2001/ast17apr_1.htm?list118443

[2] Archaefructus liaoningensis Sun,
Dilcher, Zheng et Zhou (Sun et al.,
1998). Fruiting axes and remains of two
subtending leaves (Photo courtesy of
David Dilcher). COPYRIGHTED EDU
source: http://www.flmnh.ufl.edu/deeptim
e/virtualfossilcollection/Archaeofructus
.html
mybn), end Jurassic period (206-144
mybn).
[1] Alvarezsaurid. COPYRIGHTED
source: http://www.palaeos.com/Vertebrat
es/Units/350Aves/350.200.html
evolve.

[1] Iberomesornis
COPYRIGHTED, Iberomesornis
COPYRIGHTED
source: http://www.dinosauromorpha.de/th
{kind=link}
eropoda/iberomesornis.JPG

source: http://www.lemanlake.com/photos/
{kind=link}
biotope/biodiversite/iberomesornis.gif
evolve.

[1] Sinornis santensis Artist: James
Reece COPYRIGHTED AUSTRALIA
source: http://www.amonline.net.au/chine
se_dinosaurs/feathered_dinosaurs/photo07
.htm

[1] fossil specimen of Chaoyangornis
COPYRIGHTED
source: http://www.sino-collector.com/en
{kind=link}
g/_private/cjyd/zjlt/hjs-hs/pic-l/hs0016
.jpg

[2] Chaoyangia, modified from Hou et
al. (1996) COPYRIGHTED EDU
source: http://rainbow.ldeo.columbia.edu
/courses/v1001/clover16.html
evolve.

[1] Hesperornis. COPYRIGHTED
source: http://www.savageancientseas.com
{kind=link}
/images/labels/hesperornis.jpg

[2] Detail of a painting by Ely Kish,
Copyright © Ely Kish; used with
permission of Ely Kish (EMAIL)
Hesperornis regalis Hesperornis
(pronounced HES-per-OR-nis) means
''western bird''. Toothed marine birds
of the Late Cretaceous
seas COPYRIGHTED
source: http://www.oceansofkansas.com/He
{kind=link}
sperornis/kish-01.jpg
Angiosperm group "Monocotyledons"
(Monocots) evolving now. Monocots are
the second largest lineage of flowers
after the Eudicots, and include lilies,
palms, orchids, and grasses.

[1] Sweet Flag (Acorus calamus) -
spadix Spadix of Sweet Flag. usgs
public domain
source: http://en.wikipedia.org/wiki/Aco
rus

[2] Ivy Duckweed (Lemna
trisulca) Name Lemna
trisulca Family Lemnaceae
source: http://en.wikipedia.org/wiki/Ali
smatales
Angiosperm group Eudicots (includes
most former dicotyledons) evolving now.
Eudicots are the largest lineage of
flowers.
Angiosperm groups "Asterids" and
"Rosids" evolving and separating now.
Angiosperm "Monocotyledon" (Monocot)
group "Commelinids" evolving now.

[1] Manila dwarf coconut palm from
http://www.ars.usda.gov/is/graphics/phot
os/ Manila dwarf coconut palm
thumbnail A Manila dwarf coconut palm
on the grounds of the Tropical
Agriculture Research Station in
Mayaguez, Puerto Rico. dept of
ag public domain
source: http://en.wikipedia.org/wiki/Are
cales

[2] coconut GOV public domain
source: http://www.nps.gov/kaho/KAHOckLs
{kind=link}
/KAHOplnt/images/IMG_03957.jpg
Angiosperm "Core Eudicots" evolving
now.

[1] Carnation in flower Beschreibung:
Gartennelke (Dianthus caryophyllus)
creative commons
source: http://en.wikipedia.org/wiki/Car
nation

[2] Beets GNU
source: http://en.wikipedia.org/wiki/Bee
t
brachiosaur (sauropod) fossil.
titanosaur (sauropod) fossil.
aardvarks) evolve.
meat-eating (theropod) dinosaur from
South America.
evolve.

[1] Phylum : Chordata - Class : Aves -
Order : Tinamiformes - Family :
Tinamidae - Species : Crypturellus
tataupa (Tataupa tinamou) Given to the
wikipedia by the owner, Marcos
Massarioli. Status GNU
source: http://pt.wikipedia.org/wiki/Ima
{kind=link}
gem:Crypturellus_tataupa.JPG
kiwis) evolve.
largest meat-eating dinosaur, estimated
to have been 45 to 50 feet long.
Armadillos) evolve.
Turkey, Pheasants, Peacocks, Quail)
evolve.
major line of mammals that include:
bats, camels, pigs, deer, sheep,
hippos, whales, horses, rhinos, cats,
dogs, bears, seals, walrus).
are common.
(Laurasuatheres) evolve.
shield-headed (ceratopsian) dinosaur
fossil.
(American Marsupials) evolve.
Whales (Laurasuatheres) evolve.
Marsupials) evolve.
(Laurasuatheres) evolve.
dinosaurs are common.
Hyenas, Seals, Walruses
(Laurasuatheres) evolve.
a major line of mammals that includes
rats, squirrels, rabbits, lemurs,
monkeys, apes, and humans.
Marsupials) evolve.
dinosaurs of all time exist.
Tyrannosaurus rex is the top predator
in North America and Giganotosaurus is
in South America.
club tails) evolve.
(Afrotheres) evolve.
evolve.
and pikas evolve.
Colugos evolves.
event happens.
[1] Timeline of mass extinctions.
COPYRIGHTED Benjamin Cummings.
source: http://io.uwinnipeg.ca/~simmons/
16cm05/1116/16macro.htm

[2] Cretaceous meteor impact.
COPYRIGHTED Benjamin Cummings.
source: http://io.uwinnipeg.ca/~simmons/
16cm05/1116/16macro.htm
Era.
mybn), end Cretaceous period (144-65
mybn).
evolve.
fossil mammals after the extinction of
the dinosaurs.
evolve.
marsupials) evolve.
(Australian marsupials) evolve.
marsupial) evolves.
Koalas (Australian marsupials) evolve.
gerbils, voloes, lemmings, and hamsters
evolves.
Pocket gophers, Pocket mice and
kangaroo rats evolves.
ancestor of camels and llamas splits
with the ancestor of the rest of the
Even-Toed Ungulates
(Cetardiodactyla/Artiodactyla: pigs,
ruminants, hippos, dolphins and
whales).
[1] Fig. 2. Molecular time scale for
the orders of placental mammals based
on the 16,397-bp data set and maximum
likelihood tree of ref. 14 with an
opossum outgroup (data not shown), 13
fossil constraints (Materials and
Methods), and a mean prior of 105 mya
for the placental root. Ordinal
designations are listed above the
branches. Orange and green lines denote
orders with basal diversification
before or after the K/T boundary,
respectively. Black lines depict orders
for which only one taxon was available.
Asterisks denote placental taxa
included in the ''K/T body size'' taxon
set. The composition of chimeric taxa,
including caniform, caviomorph,
strepsirrhine, and sirenian, is
indicated elsewhere (14). Numbers for
internal nodes are cross-referenced in
the supporting information.
COPYRIGHTED
source: http://www.pnas.org/content/vol1
{kind=link}
00/issue3/images/large/pq0334222002.jpeg
and Scaly-tailed Squirrels evolves.
begin to form.
Morocco.
mammal yet found, Andrewsarchus skull
dates from now {verify}.
the line that leads to the Ruminants
(cattle, goats, sheep, giraffes, bison,
buffalo, deer, wildebeast, antelope),
hippos, dolphins, and whales.
[1] Fig. 2. Molecular time scale for
the orders of placental mammals based
on the 16,397-bp data set and maximum
likelihood tree of ref. 14 with an
opossum outgroup (data not shown), 13
fossil constraints (Materials and
Methods), and a mean prior of 105 mya
for the placental root. Ordinal
designations are listed above the
branches. Orange and green lines denote
orders with basal diversification
before or after the K/T boundary,
respectively. Black lines depict orders
for which only one taxon was available.
Asterisks denote placental taxa
included in the ''K/T body size'' taxon
set. The composition of chimeric taxa,
including caniform, caviomorph,
strepsirrhine, and sirenian, is
indicated elsewhere (14). Numbers for
internal nodes are cross-referenced in
the supporting information.
COPYRIGHTED
source: http://www.pnas.org/content/vol1
00/issue3/images/large/pq0334222002.jpeg
evolve.
Mountain Beaver, Squirrels and Marmots
evolves.
Hippos split.
[1] Fig. 2. Molecular time scale for
the orders of placental mammals based
on the 16,397-bp data set and maximum
likelihood tree of ref. 14 with an
opossum outgroup (data not shown), 13
fossil constraints (Materials and
Methods), and a mean prior of 105 mya
for the placental root. Ordinal
designations are listed above the
branches. Orange and green lines denote
orders with basal diversification
before or after the K/T boundary,
respectively. Black lines depict orders
for which only one taxon was available.
Asterisks denote placental taxa
included in the ''K/T body size'' taxon
set. The composition of chimeric taxa,
including caniform, caviomorph,
strepsirrhine, and sirenian, is
indicated elsewhere (14). Numbers for
internal nodes are cross-referenced in
the supporting information.
COPYRIGHTED
source: http://www.pnas.org/content/vol1
00/issue3/images/large/pq0334222002.jpeg
[1] Figure 3: Strict consensus of 33
equally parsimonious trees with the
optimization of activity patterns.
COPYRIGHTED
source: http://www.nature.com/nature/jou
rnal/v427/n6969/fig_tab/nature02126_F3.h
tml
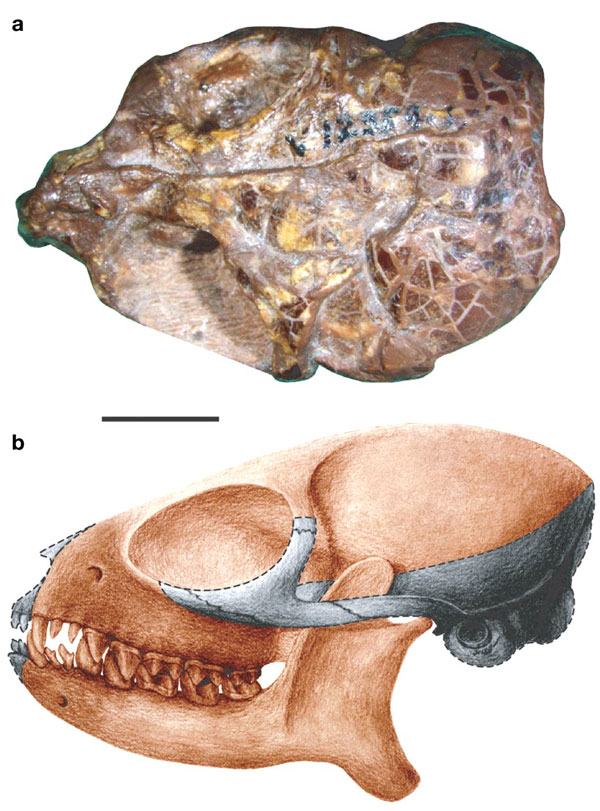
[2] FIGURE 1. The skull of Teilhardina
asiatica sp. nov. (IVPP V12357). a,
Dorsal view of the skull. b,
Reconstruction of the skull based on
IVPP V12357, with grey shadow
indicating the missing parts. Scale
bar, 5 mm. COPYRIGHTED
source: http://www.nature.com/nature/jou
rnal/v427/n6969/fig_tab/nature02126_F1.h
tml
the line to dolphins and whales split.
[1] Fig. 2. Molecular time scale for
the orders of placental mammals based
on the 16,397-bp data set and maximum
likelihood tree of ref. 14 with an
opossum outgroup (data not shown), 13
fossil constraints (Materials and
Methods), and a mean prior of 105 mya
for the placental root. Ordinal
designations are listed above the
branches. Orange and green lines denote
orders with basal diversification
before or after the K/T boundary,
respectively. Black lines depict orders
for which only one taxon was available.
Asterisks denote placental taxa
included in the ''K/T body size'' taxon
set. The composition of chimeric taxa,
including caniform, caviomorph,
strepsirrhine, and sirenian, is
indicated elsewhere (14). Numbers for
internal nodes are cross-referenced in
the supporting information.
COPYRIGHTED
source: http://www.pnas.org/content/vol1
00/issue3/images/large/pq0334222002.jpeg
whales semiaquatic "Pakicetus".
[1] Fig. 2. Molecular time scale for
the orders of placental mammals based
on the 16,397-bp data set and maximum
likelihood tree of ref. 14 with an
opossum outgroup (data not shown), 13
fossil constraints (Materials and
Methods), and a mean prior of 105 mya
for the placental root. Ordinal
designations are listed above the
branches. Orange and green lines denote
orders with basal diversification
before or after the K/T boundary,
respectively. Black lines depict orders
for which only one taxon was available.
Asterisks denote placental taxa
included in the ''K/T body size'' taxon
set. The composition of chimeric taxa,
including caniform, caviomorph,
strepsirrhine, and sirenian, is
indicated elsewhere (14). Numbers for
internal nodes are cross-referenced in
the supporting information.
. COPYRIGHTED
source: http://www.pnas.org/content/vol1
00/issue3/images/large/pq0334222002.jpeg
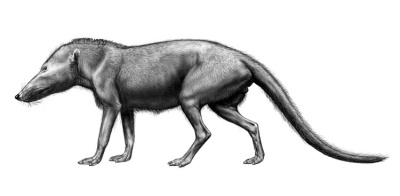
[2] Illustration by Carl Buell, and
taken from
http://www.neoucom.edu/DEPTS/ANAT/Pakice
tid.html This image is copyrighted.
The copyright holder allows anyone to
use it for any purpose, provided that
this statement is added to its caption:
''Illustration by Carl Buell, and taken
from
http://www.neoucom.edu/Depts/Anat/Pakice
tid.html ''
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Pakicetus.jpg
as India collides with Eurasia.
Pottos evolve.
eagles, Old World vultures) evolve.
dassle rats evolve.
agoutis, capybara evolve.
form.
weasel-like ancestor of bears and dogs.
Monkeys evolves.
evolve.
roadrunners, possibly hoatzin) evolve.
evolve.
whale, killer whale) and Baleen whales
(blue, humpback, gray whale) lines
split.
[1] Fig. 2. Molecular time scale for
the orders of placental mammals based
on the 16,397-bp data set and maximum
likelihood tree of ref. 14 with an
opossum outgroup (data not shown), 13
fossil constraints (Materials and
Methods), and a mean prior of 105 mya
for the placental root. Ordinal
designations are listed above the
branches. Orange and green lines denote
orders with basal diversification
before or after the K/T boundary,
respectively. Black lines depict orders
for which only one taxon was available.
Asterisks denote placental taxa
included in the ''K/T body size'' taxon
set. The composition of chimeric taxa,
including caniform, caviomorph,
strepsirrhine, and sirenian, is
indicated elsewhere (14). Numbers for
internal nodes are cross-referenced in
the supporting information.
. COPYRIGHTED
source: http://www.pnas.org/content/vol1
00/issue3/images/large/pq0334222002.jpeg
and is the largest land mammal in the
history of earth.
evolve. This Order includes many
common birds: crow, jay, sparrow,
warbler, mockingbird, robin, orioles,
bluebirds, vireos, larks, finches.
*see Toothed and baleen split.
[1] Fig. 2. Molecular time scale for
the orders of placental mammals based
on the 16,397-bp data set and maximum
likelihood tree of ref. 14 with an
opossum outgroup (data not shown), 13
fossil constraints (Materials and
Methods), and a mean prior of 105 mya
for the placental root. Ordinal
designations are listed above the
branches. Orange and green lines denote
orders with basal diversification
before or after the K/T boundary,
respectively. Black lines depict orders
for which only one taxon was available.
Asterisks denote placental taxa
included in the ''K/T body size'' taxon
set. The composition of chimeric taxa,
including caniform, caviomorph,
strepsirrhine, and sirenian, is
indicated elsewhere (14). Numbers for
internal nodes are cross-referenced in
the supporting information.
COPYRIGHTED
source: http://www.pnas.org/content/vol1
00/issue3/images/large/pq0334222002.jpeg
evolve.
evolve.
Old World Monkeys and hominids (lesser
and great apes) split.

[1] Figure 2. A synthetic hypothesis of
catarrhine primate evolution. The
branching order shown for the living
species is well-supported by numerous
molecular phylogenetic studies (for
example [6, 7, 8, 9, 10, 11, 12, 18, 24
and 25]). We present the dates of
divergence calculated by Goodman and
colleagues [11], on the understanding
that these are still rough estimates
and more precise measurements are
needed, especially for the Old World
monkeys. The fossil species (genus
names in italics) were placed on this
tree by parsimony analyses of
relatively large morphological datasets
[4, 11, 14 and 15]. Known dates for
fossils [1, 2 and 21] are indicated by
the thicker lines; these lines are
attached to the tree as determined by
the parsimony analyses, although the
dates of the attachment points are our
best guesses. Species found in Africa
are in red and species found in Eurasia
are in black. The continental locations
of the ancestral lineages were inferred
by parsimony using the computer program
MacClade [30]. The intercontinental
dispersal events required, at a
minimum, to explain the distribution of
the living and fossil species are
indicated by the arrows. COPYRIGHTED
source: http://www.sciencedirect.com/sci
ence?_ob=ArticleURL&_udi=B6VRT-4C4DVM4-D
&_user=4422&_handle=V-WA-A-W-WC-MsSAYVW-
UUW-U-AAVECYCCBC-AAVDAZZBBC-YCACYAZCV-WC
-U&_fmt=full&_coverDate=07%2F30%2F1998&_
rdoc=12&_orig=browse&_srch=%23toc%236243
%231998%23999919983%23494082!&_cdi=6243&
view=c&_acct=C000059600&_version=1&_urlV
ersion=0&_userid=4422&md5=5558415c4ccd34
6c64e2e6be03c3865e
loses tail.
evolve.
World Monkeys) evolve.
(New World Monkeys) evolve.
World Monkeys) evolve.
Monkeys) evolve.
Monkeys) evolve.
(Lesser and Great Apes), moves over
land from Africa into Europe and Asia.
[1] Figure 2. A synthetic hypothesis of
catarrhine primate evolution. The
branching order shown for the living
species is well-supported by numerous
molecular phylogenetic studies (for
example [6, 7, 8, 9, 10, 11, 12, 18, 24
and 25]). We present the dates of
divergence calculated by Goodman and
colleagues [11], on the understanding
that these are still rough estimates
and more precise measurements are
needed, especially for the Old World
monkeys. The fossil species (genus
names in italics) were placed on this
tree by parsimony analyses of
relatively large morphological datasets
[4, 11, 14 and 15]. Known dates for
fossils [1, 2 and 21] are indicated by
the thicker lines; these lines are
attached to the tree as determined by
the parsimony analyses, although the
dates of the attachment points are our
best guesses. Species found in Africa
are in red and species found in Eurasia
are in black. The continental locations
of the ancestral lineages were inferred
by parsimony using the computer program
MacClade [30]. The intercontinental
dispersal events required, at a
minimum, to explain the distribution of
the living and fossil species are
indicated by the arrows. COPYRIGHTED
source: http://www.sciencedirect.com/sci
ence?_ob=ArticleURL&_udi=B6VRT-4C4DVM4-D
&_user=4422&_handle=V-WA-A-W-WC-MsSAYVW-
UUW-U-AAVECYCCBC-AAVDAZZBBC-YCACYAZCV-WC
-U&_fmt=full&_coverDate=07%2F30%2F1998&_
rdoc=12&_orig=browse&_srch=%23toc%236243
%231998%23999919983%23494082!&_cdi=6243&
view=c&_acct=C000059600&_version=1&_urlV
ersion=0&_userid=4422&md5=5558415c4ccd34
6c64e2e6be03c3865e
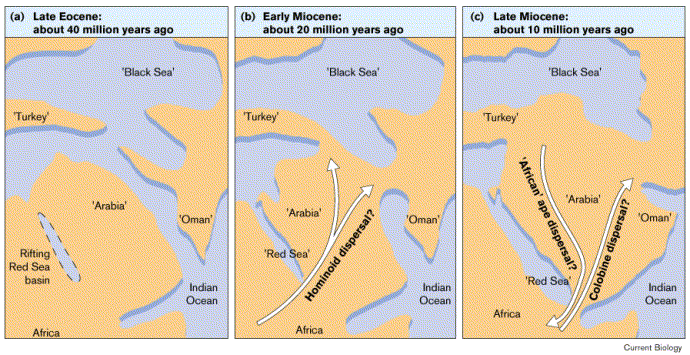
[2] Figure 1. Potential contacts
between Africa and Eurasia during the
past 40 million years, based upon
geological and faunal evidence (after
[28 and 29]). (a) Late Eocene,
approximately 40 million years ago. The
Tethys seaway prevents migration
between Africa and Eurasia. Uplifting
in the western region of the Arabian
peninsula coincides with the rifting of
the future Red Sea. (b) Early Miocene,
approximately 20 million years ago. The
Red Sea begins to form, while potential
land bridges exist between Africa and
Eurasia. (c) Late Miocene,
approximately 10 million years ago. The
Red Sea continues to grow, and
potential connections between Africa
and Eurasia exist along the Indian
Ocean margin. COPYRIGHTED
source: http://www.sciencedirect.com/sci
ence?_ob=ArticleURL&_udi=B6VRT-4C4DVM4-D
&_user=4422&_handle=V-WA-A-W-WC-MsSAYVW-
UUW-U-AAVECYCCBC-AAVDAZZBBC-YCACYAZCV-WC
-U&_fmt=full&_coverDate=07%2F30%2F1998&_
rdoc=12&_orig=browse&_srch=%23toc%236243
%231998%23999919983%23494082!&_cdi=6243&
view=c&_acct=C000059600&_version=1&_urlV
ersion=0&_userid=4422&md5=5558415c4ccd34
6c64e2e6be03c3865e
Ape Hominids) evolves in Eurasia.
[1] Figure 2. A synthetic hypothesis of
catarrhine primate evolution. The
branching order shown for the living
species is well-supported by numerous
molecular phylogenetic studies (for
example [6, 7, 8, 9, 10, 11, 12, 18, 24
and 25]). We present the dates of
divergence calculated by Goodman and
colleagues [11], on the understanding
that these are still rough estimates
and more precise measurements are
needed, especially for the Old World
monkeys. The fossil species (genus
names in italics) were placed on this
tree by parsimony analyses of
relatively large morphological datasets
[4, 11, 14 and 15]. Known dates for
fossils [1, 2 and 21] are indicated by
the thicker lines; these lines are
attached to the tree as determined by
the parsimony analyses, although the
dates of the attachment points are our
best guesses. Species found in Africa
are in red and species found in Eurasia
are in black. The continental locations
of the ancestral lineages were inferred
by parsimony using the computer program
MacClade [30]. The intercontinental
dispersal events required, at a
minimum, to explain the distribution of
the living and fossil species are
indicated by the arrows. COPYRIGHTED
source: http://www.sciencedirect.com/sci
ence?_ob=ArticleURL&_udi=B6VRT-4C4DVM4-D
&_user=4422&_handle=V-WA-A-W-WC-MsSAYVW-
UUW-U-AAVECYCCBC-AAVDAZZBBC-YCACYAZCV-WC
-U&_fmt=full&_coverDate=07%2F30%2F1998&_
rdoc=12&_orig=browse&_srch=%23toc%236243
%231998%23999919983%23494082!&_cdi=6243&
view=c&_acct=C000059600&_version=1&_urlV
ersion=0&_userid=4422&md5=5558415c4ccd34
6c64e2e6be03c3865e
[2] Figure 1. Potential contacts
between Africa and Eurasia during the
past 40 million years, based upon
geological and faunal evidence (after
[28 and 29]). (a) Late Eocene,
approximately 40 million years ago. The
Tethys seaway prevents migration
between Africa and Eurasia. Uplifting
in the western region of the Arabian
peninsula coincides with the rifting of
the future Red Sea. (b) Early Miocene,
approximately 20 million years ago. The
Red Sea begins to form, while potential
land bridges exist between Africa and
Eurasia. (c) Late Miocene,
approximately 10 million years ago. The
Red Sea continues to grow, and
potential connections between Africa
and Eurasia exist along the Indian
Ocean margin. COPYRIGHTED
source: http://www.sciencedirect.com/sci
ence?_ob=ArticleURL&_udi=B6VRT-4C4DVM4-D
&_user=4422&_handle=V-WA-A-W-WC-MsSAYVW-
UUW-U-AAVECYCCBC-AAVDAZZBBC-YCACYAZCV-WC
-U&_fmt=full&_coverDate=07%2F30%2F1998&_
rdoc=12&_orig=browse&_srch=%23toc%236243
%231998%23999919983%23494082!&_cdi=6243&
view=c&_acct=C000059600&_version=1&_urlV
ersion=0&_userid=4422&md5=5558415c4ccd34
6c64e2e6be03c3865e
divides into Cercopithecinae (Macaques
and Baboons) and Colobinae (Colobus and
Proboscis monkies).
evolve.
World Monkey) evolve.
World Monkey) evolve.
Chimpanzee, and archaic humans moves
over land from Eurasia back into
Africa.
[1] Figure 2. A synthetic hypothesis of
catarrhine primate evolution. The
branching order shown for the living
species is well-supported by numerous
molecular phylogenetic studies (for
example [6, 7, 8, 9, 10, 11, 12, 18, 24
and 25]). We present the dates of
divergence calculated by Goodman and
colleagues [11], on the understanding
that these are still rough estimates
and more precise measurements are
needed, especially for the Old World
monkeys. The fossil species (genus
names in italics) were placed on this
tree by parsimony analyses of
relatively large morphological datasets
[4, 11, 14 and 15]. Known dates for
fossils [1, 2 and 21] are indicated by
the thicker lines; these lines are
attached to the tree as determined by
the parsimony analyses, although the
dates of the attachment points are our
best guesses. Species found in Africa
are in red and species found in Eurasia
are in black. The continental locations
of the ancestral lineages were inferred
by parsimony using the computer program
MacClade [30]. The intercontinental
dispersal events required, at a
minimum, to explain the distribution of
the living and fossil species are
indicated by the arrows. COPYRIGHTED
source: http://www.sciencedirect.com/sci
ence?_ob=ArticleURL&_udi=B6VRT-4C4DVM4-D
&_user=4422&_handle=V-WA-A-W-WC-MsSAYVW-
UUW-U-AAVECYCCBC-AAVDAZZBBC-YCACYAZCV-WC
-U&_fmt=full&_coverDate=07%2F30%2F1998&_
rdoc=12&_orig=browse&_srch=%23toc%236243
%231998%23999919983%23494082!&_cdi=6243&
view=c&_acct=C000059600&_version=1&_urlV
ersion=0&_userid=4422&md5=5558415c4ccd34
6c64e2e6be03c3865e
[2] Figure 1. Potential contacts
between Africa and Eurasia during the
past 40 million years, based upon
geological and faunal evidence (after
[28 and 29]). (a) Late Eocene,
approximately 40 million years ago. The
Tethys seaway prevents migration
between Africa and Eurasia. Uplifting
in the western region of the Arabian
peninsula coincides with the rifting of
the future Red Sea. (b) Early Miocene,
approximately 20 million years ago. The
Red Sea begins to form, while potential
land bridges exist between Africa and
Eurasia. (c) Late Miocene,
approximately 10 million years ago. The
Red Sea continues to grow, and
potential connections between Africa
and Eurasia exist along the Indian
Ocean margin. COPYRIGHTED
source: http://www.sciencedirect.com/sci
ence?_ob=ArticleURL&_udi=B6VRT-4C4DVM4-D
&_user=4422&_handle=V-WA-A-W-WC-MsSAYVW-
UUW-U-AAVECYCCBC-AAVDAZZBBC-YCACYAZCV-WC
-U&_fmt=full&_coverDate=07%2F30%2F1998&_
rdoc=12&_orig=browse&_srch=%23toc%236243
%231998%23999919983%23494082!&_cdi=6243&
view=c&_acct=C000059600&_version=1&_urlV
ersion=0&_userid=4422&md5=5558415c4ccd34
6c64e2e6be03c3865e
human lives in Africa.
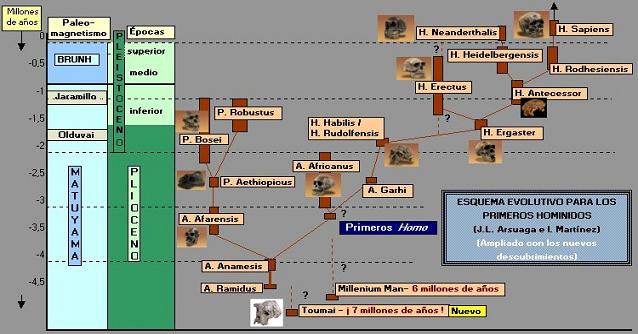
[1] evolution of the first
hominids COPYRIGHTED
source: http://www.portalciencia.net/ant
roevoerga.html
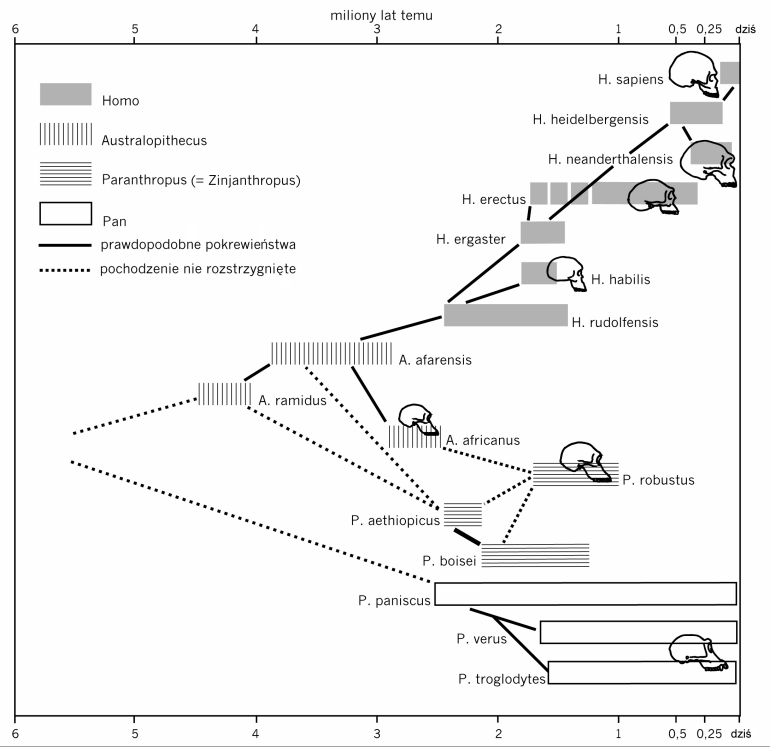
[2] Drzewo rodowe
człowiekowatych
(hominidów). hominid
evolution COPYRIGHTED
source: http://www.wiw.pl/Biologia/Ewolu
cjonizm/EwolucjaCzlowieka/BigImage.asp?c
p=1&ce=2
evolves in early hominids.
[1] evolution of the first
hominids COPYRIGHTED
source: http://www.portalciencia.net/ant
roevoerga.html
[2] Drzewo rodowe człowiekowatych
(hominidów). hominid
evolution COPYRIGHTED
source: http://www.wiw.pl/Biologia/Ewolu
cjonizm/EwolucjaCzlowieka/BigImage.asp?c
p=1&ce=2

[1] Australopithecus squinted at the
blue African sky. He had never seen a
star in broad daylight before, but he
could see one today. White. Piercing.
Not as bright as the Sun, yet much more
than a full moon. Was it dangerous? He
stared for a long time, puzzled, but
nothing happened, and after a while he
strode across the savanna
unconcerned. Millions of years
later, we know better. ''That star
was a supernova, one of many that
exploded in our part of the galaxy
during the past 10 million years,''
says astronomer Mark Hurwitz of the
University of
California-Berkeley. Right: Human
ancestors, unconcerned by odd lights in
the daytime sky. This image is based on
a painting featured in The
Economist. PD
source: http://science.nasa.gov/headline
s/y2003/06jan_bubble.htm?list847478

[2] Image Source *
http://www.familie-rebmann.de/photo11.ht
m COPYRIGHTED CLAIMED FAIR USE
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Laetoliafar.jpg.jpg
Africa.
bipedal early homonids evolves in
Africa.

[1] Skull of Paranthropus boisei. From
Smithsonian Institute
website. COPYRIGHTED CLAIMED FAIR USE
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Zinj3.jpg
[2] evolution of the first
hominids COPYRIGHTED
source: http://www.portalciencia.net/ant
roevoerga.html
This
is the earliest member of the genus
Homo.
This is when the human brain
begins to get bigger.
Homo habilis is
thought to be the ancestor of Homo
ergaster.
Homo Habilis evolved in
Africa.

[1] KNM ER 1813 Homo habilis This
image is from the website of the
Smithsonian Institution [1] and may be
copyrighted. The Smithsonian
Institution explicitly considers the
use of its content for non-commercial
educational purposes to qualify as fair
use under United States copyright law,
if: 1. The author and source of the
content is clearly cited. 2. Any
additional copyright information about
the photograph from the Smithsonian
Institution website is included. 3.
None of the content is modified or
altered.
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:KNM_ER_1813.jpg
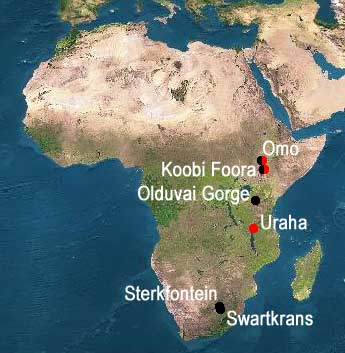
[2] red= Homo rudolfensis black=Homo
habilis COPYRIGHTED
source: http://sesha.net/eden/Eerste_men
sen.asp
thinner and less body hair.
SCIENCE
Europe and Asia. Ergaster is the first
hominid to leave Africa.
mybn-now), end Tertiary period (65-1.8
mybn).
of Africa. Homo Erectus evolves
from Homo Ergaster in Asia.
start Quaternary period (1.8 mybn-now).
of fire, from Israel.

[1] Fig. 2. Cross section of burned
Olea europaea subsp. oleaster (wild
olive) specimen. Wood is diffuse
porous; vessels are solitary and in
short radial multiples. Bar, 0.5
mm COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/304/5671/725/FIG2

[2] Fig. 3. Burned grain of Aegilops
cf. geniculata: dorsal view of a basal
fragment (this grain is also shown in
fig. S2). Parts of husk and embryo are
clearly seen. Bar, 1 mm. COPYRIGHTED
source: http://www.sciencemag.org/cgi/co
ntent/full/304/5671/725/FIG3
Africa.
transition from the verbal language of
chimps and monkeys, that will result in
the short staccato language humans use
now.
Either the majority of the 50 basic
sounds were learned simulateneously for
all sapiens by word of mouth or those
50 basic sounds evolved before the
sapiens dispersed throughout eurasia.
Since sapiens spread out over Europe
and Asia did not develop one language
with the same sounds used for each
word, it seems unlikely that the 50
basic sounds that are found in all of
those languages would not be unified
for all sapiens, and that more likely
the majority of those sounds evolved in
a smaller group in Africa and were then
dispersed into Europe, Asia, and then
Australia and the Americas.
in Ethiopia, Africa.
ergaster in Europe and Western Asia.
Oldest Neanderthal fossil in Croatia.
[93000 BCE]
Africa.
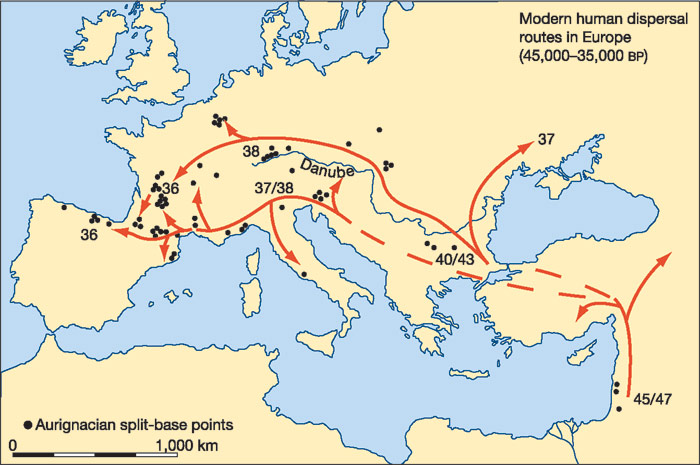
[1] The northern route (along the
Danube) is represented by the 'classic'
Aurignacian technologies, while the
southern (Mediterranean) route is
represented by the 'proto-Aurignacian'
bladelet technologies (Fig. 3)-with
their inferred origins in the preceding
early Upper Palaeolithic technologies
in the Near East and southeastern
Europe. Dates (in thousands of years
bp) indicate the earliest radiocarbon
dates for these technologies in
different areas, expressed in thousands
of radiocarbon years before present
(bp). (These are likely to
underestimate the true (calendar) ages
of the sites by between 2,000 and 4,000
yr; see ref. 32). Dashed lines indicate
uncertain routes. COPYRIGHTED
source: http://www.nature.com/nature/jou
rnal/v432/n7016/fig_tab/nature03103_F1.h
tml
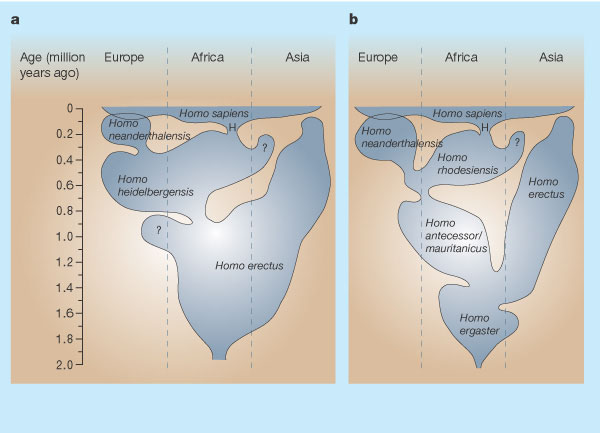
[2] The figure shows the geographical
and temporal distribution of hominid
populations, based on fossil finds,
using different taxonomic schemes. The
new finds from Herto4, 5 (H) represent
early Homo sapiens. a, This reflects
the view that both Neanderthals and
modern humans derived from a widespread
ancestral species called H.
heidelbergensis2. b, However, evidence
is growing that Neanderthal features
have deep roots in Europe2, 8, so H.
neanderthalensis might extend back over
400,000 years. The roots of H. sapiens
might be similarly deep in Africa, but
this figure represents the alternative
view that the ancestor was a separate
African species called H. rhodesiensis.
Different views of early human
evolution are also shown. Some workers
prefer to lump the earlier records
together and recognize only one
widespread species, H. erectus2 (shown
in a). Others recognize several
species, with H. ergaster and H.
antecessor (or H. mauritanicus) in the
West, and H. erectus only in the Far
East8 (shown in b). Adapted with
permission from refs 8, 11. 8.
Hublin, J.-J in Human Roots: Africa
and Asia in the Middle Pleistocene (eds
Barham, L. & Robson-Brown, K.) 99-121
(Western Academic & Specialist Press,
Bristol, 2001). 11. Rightmire, G. P.
in Human Roots: Africa and Asia in the
Middle Pleistocene (eds Barham, L. &
Robson-Brown, K.) 123-133 (Western
Academic & Specialist Press, Bristol,
2001). COPYRIGHTED
source: http://www.nature.com/nature/jou
rnal/v423/n6941/fig_tab/423692a_F1.html
[90000 BCE]
outside Africa, in Israel.
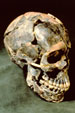
[1] Figure 2: Three-quarter view of the
Mousterian cranium Qafzeh 9 from Jebel
Qafzeh in Israel, about 92,000 years
old. Photo: Tsila
Sagiv/IDAM. COPYRIGHTED
source: http://www.metmuseum.org/special
/Genesis/tattersall_lecture.asp?printFla
g=1&refPage=1

[2] Qafzeh Cave COPYRIGHTED
source: http://www.hf.uio.no/iakh/forskn
ing/sarc/iakh/lithic/AmudNet/Asites2.htm
l
[51300 BCE]
Java.
[40000 BCE]
Australia.
[38000 BCE]
Europe.
[33000 BCE]
[28000 BCE]
China.
[8700 BCE]
artifact, from Northern Iraq.
[6000 BCE]
[6000 BCE]
[6000 BCE]
[6000 BCE]
[6000 BCE]
[6000 BCE]
[6000 BCE]
and killed for food. Goats (check: or
dogs?) are oldest domesticated animal.
[5000 BCE]
and casted (where?).
[4000 BCE]
iron from meteorites, in Egypt.
[3500 BCE]
mixed with tin) melted, and casted
(where?).
[3500 BCE]
of smelted iron, in Egypt.
[3500 BCE]
pottery wheel, comes from Mesopotamia.
[3500 BCE]
baked clay tablets is invented in Sumer
and replaces a clay token counting
system. These "numerical tablets"
represent the first recorded place
value number system (the position of
the number is multiplied by a base
number), a sexagesimal (base 60)
numbering system. This base 60
numbering system will be used
continuously to count time, for
astronomy, and geography, and is still
in use today.
The first writing begins
as numbers on clay tablets, some also
with stamped seals.
This system of
writing on clay tablets will evolve
into modern written language. Writing
was first used to solve simple
accounting problems; for example to
count large numbers of sheep or bales
of hay. Writing may have arisen out of
the need for arithmetic and storage of
information, but will grow to record
and perpetuate stories, myths, epics,
songs, and most of what we know about
human history.
Iran)

[1] MS 3007 NUMBERS 10 AND 5 +4 + 4
+ 4 + 5 + 3 ms3007MS on clay,
Syria/Sumer/Highland Iran, ca.
3500-3200 BC, 1 elliptical tablet,
6,7x4,4x1,9 cm, 2+1 compartments, 2 of
which with 3 columns of single numbers
as small circular
depressions. Commentary:Numerical or
counting tablets with their more
complex combination of decimal and
sexagesimal numbers are a further step
from the tallies with the simplest form
of counting in one-to-one
correspondence. They were used parallel
with the bulla-envelopes with tokens.
The commodity counted was not indicated
in the beginning, but was gradually
imbedded in the numbers system or with
a seal or a pictograph of the commodity
added, i. e. development into
ideonumerographical tablets, the
forerunners to pictographic tablets.
There are only about 260 numerical
tablets known. Most of them are found
in Iran. COPYRIGHTED
source: http://www.earth-history.com/_im
{kind=link}
ages/ms3007.jpg

[2] MS 4647 NUMBERS 3+4, POSSIBLY
REPRESENTING 3 MEASURES OF BARLEY AND 4
MEASURES OF SOME OTHER COMMODITY, IN
SEXAGESIMAL NOTATION ms4647MS on clay,
Syria/Sumer/Highland Iran, ca.
3500-3200 BC, 1 tablet, 4,4x5,0x2,3 cm,
2 lines with 3 small circular
depressions and 4 short
wedges. Numerical or counting
tablets with their more complex
combination of decimal and sexagesimal
numbers are a further step from the
tallies with the simplest form of
counting in one-to-one correspondence.
They were used parallel with the
bulla-envelopes with tokens. The
commodity counted was not indicated in
the beginning, but was gradually
imbedded in the numbers system or with
a seal or a pictograph of the commodity
added, i. e. development into
ideonumerographical tablets, the
forerunners to pictographic tablets.
There are only about 260 numerical
tablets known. Most of them are found
in Iran. Exhibited: The Norwegian
Intitute of Palaeography and Historical
Philology (PHI), Oslo,
13.10.2003- COPYRIGHTED
source: http://www.earth-history.com/_im
{kind=link}
ages/ms4647.jpg
[01/01/3300 BCE]
into the beginning of pictographic
writing. Now along with numbers on the
clay tablets are symbols that represent
the commodity (such as cows, sheep, and
cereals). These symbols represent the
earliest record of what will become the
modern alphabet. These tablets are all
economic records, used to keep a record
of objects owned or traded, and contain
no stories.
Writing begins as a method
for increasing the human memory to keep
track of the many transactions of a
city, and not for the purpose of
recording or remembering stories.
With the
beginning of writing, begins the first
systematic training and industry of
scribes and this will ultimately evolve
into the modern school system.

[1] MS 4551 Account of grain products,
bread, beer, butter oil. Sumer 32nd
century COPYRIGHTED?
source: http://www.earth-history.com/_im
{kind=link}
ages/ms4551.jpg

[2] MS2963 Account of male and
female slaves Sumer
c3300-3200BCE COPYRIGHTED?
source: http://www.earth-history.com/_im
{kind=link}
ages/ms2963.jpg
[01/01/3000 BCE]
script becomes phonetic (the sounds of
symbols are combined to form words).
This is the beginning of phonetic
written language.
[1] Pre-literate counting and
accounting MS 5067/1-8 NEOLITHIC
PLAIN COUNTING TOKENS POSSIBLY
REPRESENTING 1 MEASURE OF GRAIN, 1
ANIMAL AND 1 MAN OR 1 DAY'S LABOUR,
RESPECTIVELY ms5067/1-8Counting tokens
in clay, Syria/Sumer/Highland Iran, ca.
8000-3500 BC, 3 spheres: diam. 1,6, 1,7
and 1,9 cm , (D.S.-B 2:1); 3 discs:
diam. 1,0x0,4 cm, 1,1x0,4 cm and
1,0x0,5 cm (D.S.-B 3:1); 2
tetrahedrons: sides 1,4 cm and 1,7 cm
(D.S.-B 5:1). Exhibited: The
Norwegian Intitute of Palaeography and
Historical Philology (PHI), Oslo,
13.10.2003- COPYRIGHTED
source: http://www.earth-history.com/_im
{kind=link}
ages/ms5067.jpg

[2] MS 4631 BULLA-ENVELOPE WITH 11
PLAIN AND COMPLEX TOKENS INSIDE,
REPRESENTING AN ACCOUNT OR AGREEMENT,
TENTATIVELY OF WAGES FOR 4 DAYS' WORK,
4 MEASURES OF METAL, 1 LARGE MEASURE OF
BARLEY AND 2 SMALL MEASURES OF SOME
OTHER COMMODITY ms4631Bulla in clay,
Syria/Sumer/Highland Iran, ca.
3700-3200 BC, 1 spherical
bulla-envelope (complete), diam. ca.
6,5 cm, cylinder seal impressions of a
row of men walking left; and of a
predator attacking a deer, inside a
complete set of plain and complex
tokens: 4 tetrahedrons 0,9x1,0 cm
(D.S.-B.5:1), 4 triangles with 2
incised lines 2,0x0,9 (D.S.-B.(:14), 1
sphere diam. 1,7 cm (D.S.-B.2:2), 1
cylinder with 1 grove 2,0x0,3 cm
(D.S.-B.4:13), 1 bent paraboloid
1,3xdiam. 0,5 cm
(D.S.-B.8:14). Context: MSS 4631-4646
and 5114-5127are from the same archive.
Total number of bulla-envelopes
worldwide is ca. 165 intact and 70
fragmentary. COPYRIGHTED
source: http://www.earth-history.com/_im
{kind=link}
ages/ms4631.jpg
[2630 BCE]
doctor of recorded history designs the
first pyramid in Egypt.
[2600 BCE]
the Sumerian flood story.
[2500 BCE]
civilization Harappa and Mohenjo-Daro.
[2500 BCE]
dynasty) rule egypt.
[2345 BCE]
egypt.
[2300 BCE]
Sargon the Great, a semite human.
Sargon unites Sumer wth northern half
of mesopotama. Ruled from Agade, built
in South central Mesopotamia called
Akkad. The language used from this time
on in Mesopotamia is called "Akkadian".
[2181 BCE]
(7th and 8th families) .
[2160 BCE]
egypt (9 and 10th families).
[2134 BCE]
(11th family).
[2134 BCE]
decribed from Egyptian scribe humans as
"classic stage" of egyptin language.
This language is used until Roman rule
in 2186 BC. This language is used for
religious texts, narrative (?), poetry,
business documents. and is eventually
reserved for historical and religious
inscriptions on stone or papyrus.
This language is revived/used again in
Greco-Roman period for temple
inscriptions, in crytic/decorative
script called Ptolemaic.
[2040 BCE]
egypt (12th family).
[2000 BCE]
Valley.
[2000 BCE]
China.
[2000 BCE]
vehicles.
[2000 BCE]

[1] Ancient wooden lock and key from
Khorsabad (Much reduced) COPYRIGHTED
source: http://www.usgennet.org/usa/topi
c/preservation/science/inventions/chpt8.
htm
[1842 BCE]
alphabet. Proto-semitic alphabet made
in turquoise mines probably by Semitic
humans. This alphabet is thought to
have replaced cuneiform, and may be
root of all other alphabets.
[1500 BCE]
rice grown in china.
[1500 BCE]
(where?).
[1198 BCE]
egypt.
[1087 BCE]
humans in Thebes (family 21) rule
egypt.
[945 BCE]
rule egypt.
[850 BCE]
alphabet language from phoenician
humans. Phoenician humans are using a
variation of letters used at this time
by Semite humans in Syria-Palestine,
Canaanite writing. "Alef" (ox), "beth"
(house), "gimel" (camel), "daleth"
(door), etc. are changed to "alpha",
"beta", "gamma", "delta", etc. The
semitic alphabets Hebrew and Arabic are
descended from the Canaanite language.
[609 BCE]
Assyrian army of Ashur-uballit II and
capture Harran. Ashur-uballit, the last
Assyrian king, disappears from history.
[538 BCE]
conquer Babylonia, Egypt and all in
between. Jewish humans are allowed to
return to Jerusalem from captivity in
Babylonia, where they build a new
temple.
[470 BCE]
(᾿Αλκμα&#
8055;ων) (~500 BC Croton,
Italy - ???) is first to theorize that
the brain is the center of wisdom, and
emotions. Alcmaeon is the first human
known to dissect the bodies of humans
and other species. (check in )
Alcmaeon records the existence of the
optic nerve and the tube connecting the
ear and mouth, and distinguishes
arteries from veins.
[464 BCE]
Clazomenae/Klazomenai 75 mi north of
Miletus - ~428 BC Lampsacus now Lapseki
Turkey) introduces Ionian science of
Thales to Athens, saying that the
universe was not made by a diety, but
through the action of infinite "seeds",
which will later develop into atomic
theory under Leucippos. Anaxagoras
accurately explains the phases of the
earth moon, and both eclipses of moon
and sun in terms of their movements.
Anaxagoras says that the sun is a red
hot stone and the moon a real place
like the earth, not gods as is the
prevailing belief.
[460 BCE]
Λευκιπ`
0;ος ) (lEUKEPOS?)
(Leucippus) (~490 BC Miletus -???) is
the first person of record to support
the theory that everything is composed
entirely of various indestructable,
indivisible elements called atoms.
[460 BCE]
Agrigento), Sicily - Mount Etna (?)
~430 bc) understands that the heart is
the center of the blood vessel system.
Empedocles thinks some organisms not
adapted to life have died in the past.
Empedocles unites the 4 elements
(water, air, fire, earth) described by
earlier people into a theory of the
universe.
[450 BCE]
croton - ~385 BCE), the most recognized
of the Pythagorian school after
Pythagoras, theorizes that the earth
was not the center of the universe but
moves through space. Philolaus thinks
the earth, moon, the other planets and
sun circle a great fire in separate
spheres, and that the sun is only a
reflection of this fire. This is the
first recorded idea that the earth
moves thru space.
[430 BCE]
and atheism and brought to trial.
Pericles faces people in court in
defense of Anaxagoras, and Anaxagoras
is freed (unlike Socrates a generation
later).
[430 BCE]
Δημόκρ_
3;τος) (~460 BC Abdera,
thrace -~ 370 BC) in Abdera, elaborates
on atomic theory of his teacher
Leukippos. Demokritos thinks that the
Milky Way was a vast group of tiny
stars. Demokritos explains the motions
of atoms as based on natural laws, not
on the wants of gods or demons.
[430 BCE]
BCE Larissa (now Larisa), Thessaly)
founds a school of medicine on Cos that
is the most science based of the time.
Hippocrates will be recognized as the
father of medicine, although other
people (like Alcmaeon had practiced
healing and were students of the human
body). 50 books, called the
Hippocratic collection, are credited to
him, but are more likely collected
works of several generations of his
school, brought together in Alexandria
in 200-300 BCE. The books contain a
high order of logic, careful
observation, and good conduct.
Disease
was viewed as a physical phenomenon,
not credited to arrows of Apollo, or
possession by demons. For example,
epilepsy, was thought to be a sacred
disease, because a human appeared to be
in the grip of a god or demon, but in
this school epilepsy was described as
being caused by natural causes and
thought to be curable by physical
remedies, not by exorcism.
[404 BCE]
28 and 29) 404-378 BCE.
[399 BCE]
Σωκράτ_
1;ς) SO-Kro-TES? (~470 BC Athens -
399 BC Athens) is sentenced to death
and forced to end his own life, charged
with impiety, (failure to show due
piety toward the gods of Athens,
"asebia" greek:
ασέβει^
5;) and of corrupting Athenian youth
through his teachings.

[1] From
http://hypernews.ngdc.noaa.gov This
image is in the public domain because
its copyright has expired in the United
States and those countries with a
copyright term of life of the author
plus 100 years or less. PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Socrates.png

[2] The Death of Socrates, by
Jacques-Louis David (1787) The
two-dimensional work of art depicted in
this image is in the public domain in
the United States and in those
countries with a copyright term of life
of the author plus 100 years. This
photograph of the work is also in the
public domain in the United States (see
Bridgeman Art Library v. Corel Corp.).
PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Socratesdeath.jpg
[387 BCE]
Πλάτων,
Plátōn, "wide, broad-shouldered")
(~427BC Athens - 347 BC Athens) founds
a school in western Athens on a piece
of land once owned by a legendary Greek
human named "Academus", and so this
school comes to be called "The
Academy", and this word will eventually
generally apply to any school. The
Academy will be a center for science
and education for 900 years until 529
CE.
[378 BCE]
Εύδοξο`
2;) (~408 BC Cnidus (now Turkish coast)
- ~355 bc Cnidus) is the first Greek
human to realize that the year is not
exactly 365 days, but 6 hours more.
Egyptians were already aware of this
and Eudoxus may have gotten this idea
from Egypt. Eudoxus draws a map of
earth better than the map of Hecataeus.
Eudoxus is first greek human to try to
map stars. Eudoxus divides the sky in
to degrees of latitude and longitude, a
system that is eventually applied to
the earth.
[1] A pupil of Plato, Eudoxus
elaborated a geocentric model composed
of crystalline spheres, incorporating
the Platonic ideal of uniform circular
motion. System of 27 Spheres:
* 1 for the fixed stars * 3 each
for the Sun and Moon * 4 each for
the 5 planets Spheres within
spheres in perfect circular motion
combine to give retrograde
motions. Spheres within
Spheres (Click on the image to
view at full scale [Size: 20Kb]) 4
Spheres for each planet: * One
was aligned with the celestial poles,
turning once a day to give rising &
setting. * Second was tilted
23.5º, rotated slowly in the opposite
direction to give the usual
west-to-east drift of the planets
relative to the fixed stars. *
Third & Fourth were introduced to
produce the periodic retrograde motions
of the planets. All were in uniform
circular motion about their axes.
COPYRIGHTED EDU
source: http://www-astronomy.mps.ohio-st
ate.edu/~pogge/Ast161/Unit3/greek.html
[378 BCE]
Nectanabo II) rules egypt from 378 to
341 BCE.
[370 BCE]
(῾Ικέτη&
#962;), and fellow Pythagorean Ekfantos
(Έκφαντ
59;ς) (400 BCE) are the first to
theorize that the earth turns on its
own axis.
[366 BCE]
Αριστο`
4;έλης,
Aristotélēs) (ArESTOTeLAS?) opens
his own school in Athens, called the
Lyceum
(Λύκειο
57;, Lykeion) (lIKEoN?). Aristotle
classifies 500 species, and dissectes
nearly 50, correctly classifying
dolphins with species of the field, not
with fish. Aristotle puts forward the
first theory of gravity, claiming that
heavy objects go down and incoreectly
that light objects go up.
[357 BCE]
(Ηράκλε
53;τος) (387 BCE- 312
BCE) adopts the view of two
Pythagoreans, Hiketos and Ekfantos, in
theorizing that the earth rotates on
its own axis. Herakleitos thinks that
the planets Mercury and Venus orbit the
sun (although putting the earth at the
center of the universe). Herakleitos
speculates that the universe was
infinite, each star being a world in
itself, composed of an earth and other
planets.

[1]
Ηράκλε_
3;τος (~544 - 483
π.Χ.) COPYRIGHTED GREECE
source: http://sfr.ee.teiath.gr/historia
{kind=link}
/historia/important/html/images/Heraklit
.jpg
[332 BCE]
Egypt.
[323 BCE]
(Καλλιπ
60;ος) KAL lEP POS? (~370 BCE
Cyzicus - ~ 300 BCE) makes a more
accurate measurement of the solar year,
finding the measurement of Meton 100
years earlier to be 1/76 of a day too
long. Kallippos constructs a a 76 year
cycle of 940 months to unite the solar
and lunar years. This calendar is
adopted in 330 BCE and will be used by
all later astronomers.
[311 BCE]
(Επίκου
61;ος) (Epicurus) (02/341 BCE
Samos - 270 BCE Athens) founds a
popular school in Athens. He argues
against the existence of any god.
Epikouros basis his philosophy on the
principle that pleasure is good and
pain is bad. This is the first school
to admit females and slaves. Epikouros
agrees with the atom theory of
Demokritos.
[305 BCE]
(Ηροφιλ
59;ς) (Herophilus) (335 BCE
Chalcedon {now Kadikoy, Istanbul
Turkey} - 280 BCE) is the first human
to distinguish nerves from blood
vessels, in addition to motor nerves
from sensory nerves.
Herofilos is the first to
describe the liver and spleen, to
describe and name the retina of the
eye, to name the first section of the
small intestine "the duodenum", to
describe ovaries, the tubes leading to
the ovaries from the uterus, and names
the prostate gland. Herofilos is the
first human to note that arteries carry
blood, not air as previously believed,
a recognizes that the heart pumps blood
through the blood vessels. Herofilos is
first to distinguish between cerebrum
and cerebellum.
[295 BCE]
Εὐκλεί
48;ης) YUKlEDES? (325 BCE -
265 BCE), in Alexandria, makes a scroll
called "Elements" which is a
compilation of all the mathematical
knowledge known up to then, and will be
one of the most successful mathmatical
texts in the history of earth.
Euclid
proves that the number of primes is
infinite, that the square root of 2 is
irrational, and shows light rays as
straight lines.
[285 BCE]
(Greek
Κτησίβ_
3;ος), (fl. 285 - 222 BCE) a
member of the Alexandrian Mouseion, is
the first person of record to use
compressed air, building a water and
compressed air powered organ and
catapult.
[1] Ktesibios water organ. COPYRIGHTED
source: http://alexandrias.tripod.com/ct
esibius.htm
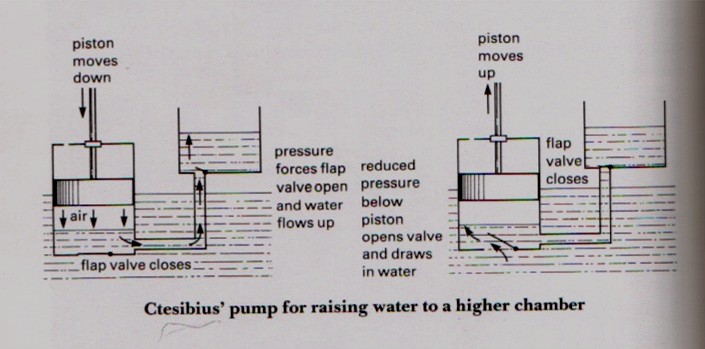
[2] Ktesibios water pump. COPYRIGHTED
source: http://alexandrias.tripod.com/ct
esibius.htm
[283 BCE]
that the earth and other planets go
around the sun. Aristarchus figures
out that the Sun is one of the fixed
stars, the closest star to the Earth.
Aristarchos understands the earth
rotates on it's own axis each day.
Aristarchos understands that the sun is
much larger than the earth.
Aristarchos understands that the stars
are very distant. Aristarchos
calculates a close estimate for the
size of the earth moon. A principle
work of Aristarchos, titled
"Heliocentric system", now lost, is
considered by many of his
contemporaries as "impious", and one
contemporary writes that Aristarchos
should be charged with impiety.
[274 BCE]
Ερασίσ`
4;ρατος
(EroSESTrATOS?) (~304 BCE Chios {now
Khios, an aegean island} - 250 BCE
Samos), in Alexandria describes the
brain as being divided in to a larger
cerebrum and smaller cerebellum.
Erasistratos accepts atom theory.
[257 BCE]
Αρχιμή^
8;ης ) (287 Syracuse, Sicily
- 212 Syracuse, Sicily) is the first to
understand density (how mass and volume
are related). Archimedes makes a system
that is equivalent to the exponential
system to describe the amount of sand
needed to fill the universe. He makes
the best estimate of pi, builds a
mechanical model of the universe, and a
"screw of Archimedes".

[1] In the process, he calculated the
oldest known example of a geometric
series with the ratio 1/4 GNU
source: http://en.wikipedia.org/wiki/Arc
himedes

[2] parabola and inscribed triangle.
PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Parabola.png
[250 BCE]
(Απολλώ
57;ιος ο
Περγαί_
9;ς ) (261 BCE Perga {south coast
of Turkey} - 190 BCE Pergamum?) is the
first to describe the ellipse,
parabola, and hyperbola.
[246 BCE]
(Ἐρατοσθένης) (276 BCE
Cyrene now Shahat, on Libyan coast -
196 BCE Alexandria) is the first person
to accurately calculate the size of the
earth.
[160 BCE]
Ἳππαρχ
59;ς) (Nicaea {now Iznik in NW
Turkey} 190 BCE - 120 BCE), astronomer
in the Mouseion in Alexandria, uses a
solar eclipse to determine the distance
from the Earth to the Moon. Hipparchos,
is the first person to make a
trigonometric table, and is probably
first to develop a reliable method to
predict solar eclipses. Hipparchos
compiles a star catalog with 850 stars
and their relative brightness, and
probably invents the astrolabe.
Hipparchos does not use the
sun-centered system of Aristarchos, but
instead the mistaken earth-centered
system Anaxamander and the vast
majority of others chose to support.

[1] image of Hipparchos from coin?
http://www-history.mcs.st-and.ac.uk/hist
ory/Mathematicians/Hipparchus.html PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Hipparchos_1.jpeg

[2] hipparchos stamp UNKNOWN
source: http://www-history.mcs.st-and.ac
.uk/history/PictDisplay/Hipparchus.html
[56 BCE]
light as being made of tiny atoms that
move very fast.
[50 CE]
Ήρων ο
Αλεξαν^
8;ρεύς) (c.10 CE -
c.70 CE), a Greek engineer in
Alexandria, makes the first recorded
steam engine.
[1] Hero's aeolipile From Knight's
American Mechanical Dictionary, 1876.
PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Aeolipile_illustration.JPG
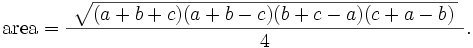
[2] Heron's formula can also be
written this way. GNU
source: http://en.wikipedia.org/wiki/Her
on%27s_formula
[77 CE]
Cecilius Secundus") (PlinE) (23 CE
Novum Comum (now Como), Italy - August
24, 79 CE near Mount Vesuvius, Italy)
completes his major work titled
"Natural History" in 37 volumes.
[391 CE]
Serapis (the Serapeum) in Alexandria is
violently destroyed by Christian people
and the temple is converted to a
church.
[416 CE]
permanently destroyed by Christian
people.
[529 CE]
the Academy in Athens.
[642 CE]
[10/12/1492 CE]
1451-1506) lands on a small island
(probably San Salvador) in America.

[1] Portrait of Christopher Columbus
from the painting Virgen de los
Navegantes (in the Sala de los
Almirantes, Royal Alcazar, Seville). A
painting by Alejo Fernández between
1505 and 1536. It is the only state
sponsored portrait of the First Admiral
of the Indias. Photo by a Columbus
historian, Manuel Rosa. More info
http://www.UnmaskingColumbus.com PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Christopher_Columbus_Face.jpg

[2] Christopher Columbus, conjectural
image by Sebastiano del Piombo in the
Gallery of Illustrious Men (Corridoio
Vasariano), Uffizi, Florence but
yet: Christophorus Columbus/Cristobal
Colon, pictue by Sebastiano del Piombo
from the XVI (15th century) PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:CristobalColon.jpg
[1609 CE]
1564-1642), understands that the
distance covered by a falling body is
proportional to the square of the
elapsed time.

[1] Galileo Galilei. Portrait in crayon
by Leoni Source: French WP
(Utilisateur:Kelson via
http://iafosun.ifsi.rm.cnr.it/~iafolla/h
ome/homegrsp.html) PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Galilee.jpg

[2] Original portrait of Galileo
Galilei by Justus Sustermans painted in
1636. PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Galileo.arp.300pix.jpg
[1609 CE]
understands that planets move in
elliptical orbits.
Stuttgart Region in the German state of
Baden-Württemberg, 30 km west of
Stuttgart's center)

[1] Johannes Kepler, oil painting by an
unknown artist, 1627; in the cathedral,
Strasbourg, France. Erich Lessing/Art
Resource, New York PD
source: http://www.britannica.com/eb/art
-2965/Johannes-Kepler-oil-painting-by-an
-unknown-artist-1627-in?articleTypeId=1

[2] A 1610 portrait of Johannes Kepler
by an unknown PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Johannes_Kepler_1610.jpg
[01/??/1610 CE]
around Jupiter and determines their
period.

[1] Galileo's Letter to Prince of
Venice PD
source: http://www2.jpl.nasa.gov/galileo
{kind=link}
/ganymede/manuscript1.jpg

[2] Galileo's illustrations of the
Moon, from his Sidereus Nuncius (1610;
The Sidereal Messenger). Courtesy of
the Joseph Regenstein Library, The
University of Chicago PD
source: http://www.britannica.com/eb/art
-2914/Galileos-illustrations-of-the-Moon
-from-his-Sidereus-Nuncius?articleTypeId
=1
[1637 CE]
(DAKoRT) describes the Cartesian
coordinate system.

[1] Portrait of René Descartes by
Frans Hals (1648) Description René
Descartes, french philosopher (Oil on
canvas, 68 x 77, Owned by the Musée du
Louvre Paris) Source No source
specified. Please edit this image
description and provide a source. Date
1648 Author Frans Hals PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Descartes.jpg

[2] Scientist: Descartes, René (1596
- 1650) Discipline(s): Physics ;
Mathematics Print Artist: William
Holl Medium: Engraving Original
Artist: Franz Hals, ca.1582-1666
Original Dimensions: Graphic: 12.7 x
10.3 cm / Sheet: 25.5 x 17.5 cm PD
source: http://www.sil.si.edu/digitalcol
lections/hst/scientific-identity/CF/disp
lay_results.cfm?alpha_sort=d
[1643 CE]

[1] Frontispiece to ''Lezioni
accademiche d'Evangelista
Torricelli....'', published in 1715.
Library Call Number Q155 .T69
1715. Image ID: libr0367, Treasures of
the NOAA Library Collection
Photographer: Archival Photograph by
Mr. Steve Nicklas, NOS, NGS Secondary
source: NOAA Central Library National
Oceanic & Atmospheric Adminstration
(NOAA), USA
http://www.photolib.noaa.gov/library/lib
r0367.htm PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Libr0367.jpg

[2] Frontispiece and title page to
''Lezioni accademiche d'Evangelista
Torricelli ....'', published in 1715.
Library Call Number Q155 .T69
1715. Image ID: libr0366, Treasures of
the NOAA Library Collection
Photographer: Archival Photograph by
Mr. Steve Nicklas, NOS, NGS Secondary
source: NOAA Central Library National
Oceanic & Atmospheric Adminstration
(NOAA),
USA http://www.photolib.noaa.gov/librar
y/libr0366.htm PD
source: http://commons.wikimedia.org/wik
{kind=link}
i/Image:Libr0366.jpg
[02/19/1672 CE]
revives the view that light is a
particle.

[1] Description Isaac Newton Date
1689 Author Godfrey Kneller PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:GodfreyKneller-IsaacNewton-1689.jpg

[2] Sir Isaac Newton Description
National Portrait Gallery
London Source
http://www.nd.edu/~dharley/HistIdeas/Ne
wton.html (not actual); first uploaded
in German Wikipedia by Dr. Manuel Date
26. Jan. 2005 (orig. upload) Author
Godfrey Kneller (1702) PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Isaac_Newton.jpeg
[1687 CE]
describes the universal law of
gravitation, that all matter attracts
other matter in a force that is the
product of their masses, and the
inverse of their distance squared.

[1] Sir Isaac Newton's own first
edition copy of his Philosophiae
Naturalis Principia Mathematica with
his handwritten corrections for the
second edition. The first edition was
published under the imprint of Samuel
Pepys who was president of the Royal
Society. By the time of the second
edition, Newton himself had become
president of the Royal Society, as
noted in his corrections. The book can
be seen in the Wren Library of Trinity
College, Cambridge. CC
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:NewtonsPrincipia.jpg
[2] Description Isaac Newton Date
1689 Author Godfrey Kneller PD
source: http://en.wikipedia.org/wiki/Ima
ge:GodfreyKneller-IsaacNewton-1689.jpg
[11/12/1801 CE]
light.
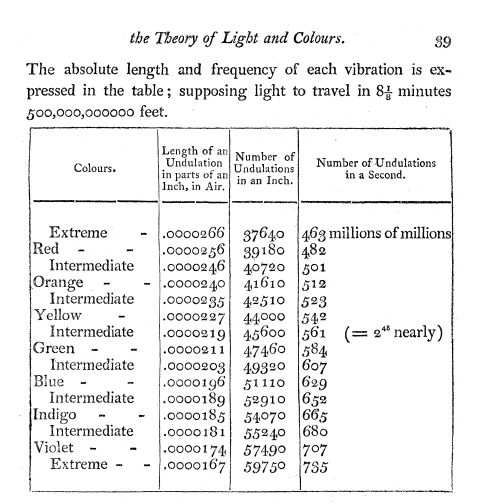
[1] [t Table of light wavelengths and
frequencies calculated by Young from
Theory of Light and Colours
11/12/1801] The inch used in the table
is the French (Paris) inch of
27.07mm. PD/Corel
source: Young_Thomas_1802_on_the_theory_
of_light_and_colours.pdf

[2]
http://journals.royalsociety.org/content
/q3r7063hh2281211/?p=422e575bae414c9a974
a16d595c628d0π=24 The Bakerian
Lecture: On the Theory of Light and
Colours Journal Philosophical
Transactions of the Royal Society of
London (1776-1886) Issue Volume 92 -
1802 Pages 12-48 DOI 10.1098/rstl.1802
.0004 Young_Thomas_1802_on_the_theory_o
f_light_and_colours.pdf [t Young
writes: ''Let the concentric lines in
Fig. 1 (Plate I.) represent the
contemporaneous situation of similar
parts of a number of successive
undulations diverging from the point A;
they will also represent the successive
situations of each individual
undulation: let the force of each
undulation be represented by the
breadth of the line, and let the cone
of light ABC be admitted through the
apeture BC; then the principal
undulations will proceed in a
recilinear direction towards GH, and
the faint radiations on each side will
diverge from B and C as centres,
without receiving any additional force
from any intermediate point D of the
undulation, on account of the
inequality of the lines DE and DF. But
if we allow some little lateral
divergence from the extremities of the
undulations, it must diminish their
force, without adding materially to
that of the dissipated light; and their
termination, instead of the right line
BG, will assume the form CH; since the
loss of force must be more considerable
near to C than at greater distances.
This line corresponds with the boundary
of the shadow in NEWTON's first
observation, Fig. 1; and it is much
more probable that such a dissipation
of light was the cause of the increase
of the shadow in that observation, than
that it was owing to the action of the
inflecting atmosphere, which must have
extended a thirtieth of an inch each
way in order to produce it; especially
when it is considered that the shadow
was not diminished by surrounding the
hair with a denser medium than air,
which must in all probability have
weakened and contracted its inflecting
atmosphere. In other circumstances, the
lateral divergence might appear to
increase, instead of diminishing, the
breadth of the
beam.''] PD/COPYRIGHTED
source: http://journals.royalsociety.org
/content/q3r7063hh2281211/?p=422e575bae4
14c9a974a16d595c628d0π=24
[10/21/1803 CE]
chemically how all matter is made of
atoms.

[1] Engraving of a painting of John
Dalton Source Frontispiece of John
Dalton and the Rise of Modern Chemistry
by Henry Roscoe Date 1895 Author
Henry Roscoe (author), William Henry
Worthington (engraver), and Joseph
Allen (painter) [t right one finger =
?] PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Dalton_John_desk.jpg

[2] John Dalton John Dalton,
1766-1844, English chemist and Fellow
of the Royal Society. [t this pose,
hand in coat=?, famous Napoleon
pose] PD/COPYRIGHTED
source: http://www.english.upenn.edu/Pro
jects/knarf/People/dalton.html
[10/24/1810 CE]
sounds heard by a brain by measuring
electricity from human nerves.

[1] 1807 engraving of camera lucida in
use Obtained from the university
website
http://www.usc.edu/schools/annenberg/asc
/projects/comm544/library/
images/448.jpg, image edited for size
and clarity. I emailed the contact at
that site and said >
http://www.usc.edu/schools/annenberg/asc
/projects/comm544/library/
images/448.jpg > is described as an
1807 picture of a camera lucida. Can
you confirm > that it isn't under
copyright? Is it OK with you if I use
it in a > Wikipedia (free Internet
encyclopedia) article on the camera
lucida? I got this
reply Daniel, This work is not
copyrighted, so far as I know--and
after 196 years, I'm quite certain any
original copyright would have long ago
expired, don't you think? Your own use
is entirely up to you--I wish you every
success. -- Jim Beniger PD
source: http://www.sil.si.edu/digitalcol
lections/hst/scientific%2Didentity/CF/di
splay_results.cfm?alpha_sort=W

[2] Optics of Wollaston camera
lucida From W. H. C. Bartlett,
Elements of Natural Philosophy, 1852,
A. S. Barnes and Company. Photocopy
kindly provided by Tom Greenslade,
Department of Physics, Kenyon College.
This image was scanned from the
photocopy and cleaned up by Daniel P.
B. Smith. This version is licensed by
Daniel P. B. Smith under the terms of
the Wikipedia Copyright. PD
source: http://www.sil.si.edu/digitalcol
lections/hst/scientific%2Didentity/CF/di
splay_results.cfm?alpha_sort=W
[10/24/1810 CE]
sounds of thought by measuring
electricity directly from human nerves.
[1811 CE]
are seen and recorded by measuring the
electricity the images produce in the
human nerves.
(add image)
[1812 CE]
thinks of are seen and recorded by
measuring the electricity the
thought-images produce in the human
nerves.
(add image)
[1812 CE]
remotely. (neuron writing)
(add image)
[1816 CE]

[1] C. Laguiche. Joseph Nicéphore
Niépce. ca1795. Ink and
watercolor. 18.5 cm in
diameter. PD/COPYRIGHTED
source: http://www.hrc.utexas.edu/exhibi
tions/permanent/wfp/3.html

[2] English: By Nicéphore Niépce in
1826, entitled ''View from the Window
at Le Gras,'' captured on 20 × 25 cm
oil-treated bitumen. Due to the 8-hour
exposure, the buildings are illuminated
by the sun from both right and left.
This photo is generally considered the
first successful permanent
photograph. PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:View_from_the_Window_at_Le_Gras%2C_Jo
seph_Nic%C3%A9phore_Ni%C3%A9pce.jpg
[04/21/1820 CE]
magnetism.

[1] A younger Hans Christian Ørsted,
painted in the 19th century. PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:%C3%98rsted.jpg

[2] Picture number :317 CD number
:9 Picture size :757x859[pixels],
66x75[mm] Date taken :0000-00-00
Date added
:2000-04-13 Fotographer/Owner :Engrave
d Location
:Denmark Description H.C. Oersted
(1777-1851). Danish physicist. Here as
a youngster. The picture was donated to
the Danish Polytech Institute,
Copenhagen, by his daughter Miss
Mathilde Oersted, April 19,
1905. PD/COPYRIGHTED
source: http://www.polytechphotos.dk/ind
ex.php?CHGLAN=2&CatID=286
[09/11/1821 CE]
England
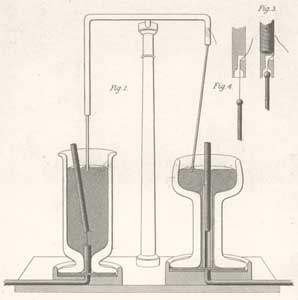
[1] The first electric motors - Michael
Faraday, 1821 From the Quarterly
Journal of Science, Vol XII, 1821 PD
source: http://www.sparkmuseum.com/MOTOR
S.HTM
[2] Description Michael Faraday,
oil, by Thomas Phillips Source
Thomas Phillips,1842 Date
1842 Author Thomas Phillips[3
wiki] The portrait shown here was
painted by Thomas Phillips (1770-1845),
oil on canvas, The National Portrait
Gallery, London.[7] PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:M_Faraday_Th_Phillips_oil_1842.jpg
[03/27/1829 CE]
magnet.

[1] Francesco Zantedeschi PD/Corel
source: http://www.liceofoscarini.it/sto
ria/bio/zantedeschi.html

[2] Image of Francesco Zantedeschi
1797 to 1873 to illustrate that
article. Uploaded from
http://www.jergym.hiedu.cz/~canovm/objev
ite/objev4/zan.htm and
http://www.jergym.hiedu.cz/~canovm/objev
ite/objev4/zan2.htm (English
translation) This portrait of
Francesco Zantedeschi was published by
Stefano de Stefani, president of the
Academy of Agriculture, Arts and
Commerce of Verona, on March 21, 1875
to accompany his eulogy to Zantedeschi
on the occasion of the transport of his
ashes to the cemetery at Verona. Black
and white version PD
source: http://en.pedia.org//Image:Franc
{kind=link}
esco_Zantedeschi_bw.jpg
[02/17/1831 CE]
England
[1] Description Michael Faraday,
oil, by Thomas Phillips Source
Thomas Phillips,1842 Date
1842 Author Thomas Phillips[3
wiki] The portrait shown here was
painted by Thomas Phillips (1770-1845),
oil on canvas, The National Portrait
Gallery, London.[7] PD
source: http://en.wikipedia.org/wiki/Ima
ge:M_Faraday_Th_Phillips_oil_1842.jpg

[2] Michael Faraday - Project
Gutenberg eText 13103 From The Project
Gutenberg eBook, Great Britain and Her
Queen, by Anne E.
Keeling http://www.gutenberg.org/etext/
13103 PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:Michael_Faraday_-_Project_Gutenberg_e
Text_13103.jpg
[09/??/1831 CE]
England
[1] Description Michael Faraday,
oil, by Thomas Phillips Source
Thomas Phillips,1842 Date
1842 Author Thomas Phillips[3
wiki] The portrait shown here was
painted by Thomas Phillips (1770-1845),
oil on canvas, The National Portrait
Gallery, London.[7] PD
source: http://en.pedia.org//Image:M_Far
{kind=link}
aday_Th_Phillips_oil_1842.jpg
[2] Michael Faraday - Project
Gutenberg eText 13103 From The Project
Gutenberg eBook, Great Britain and Her
Queen, by Anne E.
Keeling http://www.gutenberg.org/etext/
13103 PD
source: http://en.pedia.org//Image:Micha
{kind=link}
el_Faraday_-_Project_Gutenberg_eText_131
03.jpg
[04/??/1845 CE]
Ireland

[1] Abb. 2 - Lord Rosse's drwaing of M
51 showing its spiral structure. [t
Notice that Parsons numbers stars which
appear to be part of the
galaxy] PD/Corel
source: http://www.klima-luft.de/steinic
ke/Artikel/birr/birr_e.htm

[2] en: This is the sketch made by
Lord Rosse of the Whirlpool Galaxy in
1845. PD
source: http://en.wikipedia.org/wiki/Ima
{kind=link}
ge:M51Sketch.jpg
[09/23/1846 CE]
France)

[1] Scientist: Le Verrier, Urbain Jean
Joseph (1811 - 1877) Discipline(s):
Astronomy Print Artist: Auguste Bry,
19th C. Medium: Lithograph
Original Dimensions: Graphic: 12.5 x
10 cm / Sheet: 26.1 x 17 cm PD/Corel
source: http://upload.wikimedia.org/wiki
{kind=link}
pedia/commons/8/89/Urbain_Le_Verrier.jpg

[2] Scientist: Le Verrier, Urbain
Jean Joseph (1811 -
1877) Discipline(s): Astronomy Print
Artist: E. Buechner Medium:
Engraving Original Dimensions:
Graphic: 14.5 x 13 cm / Sheet: 19.5 x
14.2 cm PD/Corel
source: http://www.sil.si.edu/digitalcol
{kind=link}
lections/hst/scientific-identity/fullsiz
e/SIL14-L003-01a.jpg
[07/01/1858 CE]
from a single ancestor and the process
of natural selection.
England

[1] ''Charles Darwin, aged 51.''
Scanned from Karl Pearson, The Life,
Letters, and Labours of Francis Galton.
Photo originally from the 1859 or
1860. PD
source: http://upload.wikimedia.org/wiki
{kind=link}
pedia/commons/4/42/Charles_Darwin_aged_5
1.jpg
[2] Charles Darwin as a 7-year old boy
in 1816 The seven-year-old Charles
Darwin in 1816, one year before his
mother's death. [t A rare smile, there
are not many photos of Darwin
smiling.] PD
source: http://upload.wikimedia.org/wiki
{kind=link}
pedia/en/6/6c/Charles_Darwin_1816.jpg
[10/20/1859 CE]
spectra can be used to determine atomic
composition.
Germany
[1] Bunsen-Kirchhoff spectroscope with
the Bunsen burner (labeled D), from
Annalen der Physik (1860). Chemical
Heritage Foundation
Collections. PD/Corel
source: http://www.chemheritage.org/clas
{kind=link}
sroom/chemach/images/lgfotos/04periodic/
bunsen-kirchhoff2.jpg

[2] [t Clearly and early spectroscope,
is this from Bunsen?] PD/Corel
source: http://people.clarkson.edu/~ekat
{kind=link}
z/scientists/bunsen_spectrometer.jpg
[07/21/1969 CE]
[2015 CE]
robot assistants.
[2040 CE]
of traffic above the street level paved
roads.
[2050 CE]
replaced humans in most low-skill jobs
(fast-food, fruit and vegtable picking,
etc)
[2100 CE]
earth are aware that thought can be
seen and heard, almost 200 years after
its invention.
[2100 CE]
[2140 CE]
atoms (Silicon, Aluminum, Iron, and
Calcium) into the much more useful H2,
N2 and O2. This allows humans to live
independently of earth, on planets and
moons without water.
[2150 CE]
democratic, where the people vote
directly on the laws.
[2200 CE]
earth.
[2275 CE]
developed nations are not religious.
[2420 CE]
developed nations do not believe in any
gods.
[2500 CE]
earth orbit.
[2500 CE]
project is started.
[2500 CE]
are not religious.
[2500 CE]
[2600 CE]
uncomfortably large at 1 trillion
(1e12) humans.
[2650 CE]
particles.
[2750 CE]
different star.
[2800 CE]
not believe in any gods.
[3000 CE]
humans. Humans learn to change the
human genome in order to grow to a
certain age and maintain that age
without aging any farther. This has an
immediate impact on the population
growth of humans in the star system,
increasing the population very quickly,
limited only by water and food.
[3100 CE]
[3500 CE]
completed. Venus becomes second earth
(although without oceans and much more
efficiently organized).
[3800 CE]
reaches physical maximum of 250
trillion (250e12) humans.
[3900 CE]
Mars reaches physical maximum of 500
trillion (500e12) humans.
[4800 CE]
physical maximum of 1 quadrillion
humans (1e15).
[5000 CE]
earth reaches a theoretical physical
maximum of 333 quadrillion (333e15)
humans.
[5100 CE]
life that evolved around a different
star.
[6000 CE]
evolved around a different star.
globular galaxy.
Galaxies unite.
unite.